

Лекція 12: Клітина як цілісна система. Енергетичний обмін речовин у клітині.
МЕТАБОЛІЗМ (від греч. metabole- зміна, перетворення), 1) те ж, що обмін речовин. 2) У більш вузькому сенсі М.- проміжний обмін, що охоплює всю сукупність реакцій, головним чином ферментативних, які протікають в клітинах і забезпечують як розщеплювання складних сполук, так і їх синтез і взаємоперетворення.
Асиміляція – процеси, пов’язані з поглинанням із довкілля, засвоєнням і накопиченням хімічних речовин, які використовуються для синтезу сполук, подібних для організму, називають асиміляцією.
Енергетичний обмін – сукупність реакцій розщеплення складних сполук, які супроводжуються виділенням енергії.
Пластичний обмін – сукупність реакцій синтезу, що забезпечують ріст клітин і поновлення їхнього хімічного складу
Дисиміляція - процеси обміну речовин, які приводять до розкладу певних сполук в клітинах
Сукупність реакцій розчеплення називають енергетичним обміном клітин, або дисиміляцією. Дисиміляція прямо протилежна асиміляції: в результаті розщеплення речовини втрачають подібність до речовини клітини.
Пластичний і енергетичний обміни ( асиміляція і дисиміляція ) нерозривно зв’язані між собою. З одного боку, реакції біосинтезу потребують затрати енергії, яка черпається з реакцій розщеплення. З другого боку, для здійснення реакцій енергетичного обміну потрібний постійний біосинтез ферментів, що обслуговують ці реакції, бо в процесі своєї діяльності вони зношуються і руйнуються.
АВТОТРОФНІ ОРГАНІЗМИ, автотрофи (від авто... і ...троф), організми, що використовують для побудови свого тіла CO2 як єдине або головне. джерела вуглецю і що володіють як системою ферментів для асиміляції СОз, так і здатністю синтезувати всі компоненти клітини. Деякі А. о. можуть потребувати екзогенних (що поступають ззовні) вітамінів або чинників зростання. А. о. протиставляються ГЕТЕРОТРОФНИМ ОРГАНІЗМАМ що використовують екзогенні органічні речовини.
До А. о. відносяться:
1. наземні зелені рослини,
2. водорості,
3. фототрофні бактерії, здібні до фотосинтезу,
4. деякі бактерії що використовують окислення неорганічних речовин - хемоавтотрофи (див. Хемосинтез).
Більшість А. о. асимілює CO2 через відновний пентозофосфатний шлях. У деяких бактерій, наприклад, метанобразуючих, CO2 асимілюється іншим шляхом. Обговорюється можливість віднесення до А. о. бактерій, що використовують як джерело вуглецю метан. А. о.- первинні продуценти органічних речовини в біосфері, створюючі перший трофічний рівень у співтовариствах. Роль фотосинтезуючих А. о. в природі є таким, що визначає, оскільки вони утворюють основну масу органічної речовини в біосфері - быля. 162 • 109 т/год, в т.ч. 2/3 дають наземні рослини. Внесок інших груп А. о. кількісно невеликий. Діяльністю А. о. визначається як існування всіх інших організмів, так і хід біогеохімічних циклів в круговороті речовин в природі.
ОБМІН РЕЧОВИН, метаболізм, сукупність тих, що протікають в живих організмах хімічних перетворень що забезпечують їх зростання, життєдіяльність, відтворення, постійний контакт і обмін з навколишнім середовищем. Завдяки О. в. відбувається розщеплювання і синтез молекул, що входять в склад клітин, утворення, руйнування і оновлення клітинних структур і міжклітинної речовини. Наприклад у людини половина всіх тканинних білків розщеплюється і будується наново в середньому протягом 80 діб, білки печінки до сироватки крові наполовину оновлюються кожні 10 діб, а білки м’язів- 180, ферменты печінки - кожні 2-4 години. О. р. не можливо відокремити від процесів перетворення енергії: потенційна енергія хімічних зв'язків складних органічних молекул в результаті хімічних перетворень переходить в інші види енергії, використовуваної на синтез нових сполук, для підтримки структури і функції клітин, температури тіла, для здійснення роботи і т.д. Всі реакції О. в. і перетворення енергії протікають за участю біологічних каталізаторів - ферментів. У самих різних організмів О. в. відрізняється впорядкованістю і схожістю послідовності ферментативних перетворень не дивлячись на великий асортимент хімічних сполук, що залучаються до обміну. В той же час для кожного виду характерний особливий, генетично закріплений тип О. в., обумовлений умовами його існування. О. в. складається з двох взаємопов'язаних, що одночасно протікають в організмі процесів - асиміляції і дисиміляції, або анаболізму і катаболізму. В ході катаболічних перетворень відбувається розщеплювання великих органічних молекул до простих сполук з одночасним виділенням енергії, яка запасається у формі багатих енергією фосфатних зв'язків, головним чином. у молекулі аденозінтрифосфорної к-ти (АТФ) і інших багатих енергією з'єднань.
Катаболічні перетворення звичайно здійснюються в результаті гідролітичних і окиснювальних реакцій і протікають як у відсутність кисню (анаеробний шлях - гліколіз, бродіння), так і при його участі (аеробний шлях - дихання). В першому випадку під час гліколізу завдяки виділенню енергії відновлюється 2 молекули АТФ при рощепленні однієї молекули глюкози. Другий шлях еволюційно молодший і в енергетичному відношенні більш вигідний. Він забезпечує повне розщеплювання органічних молекул до CO2 і Н2О. Різноманітні органічні сполуки в ході катаболічних процесів перетворюються на обмежене число невеликих молекул (крім СО2 і Н2О): вуглеводи - в тріозофосфати і (або) пірувати (піровіноградна кислота), жири - в ацетіл-КоА пропіоніл-КоА і гліцерин, білки - в ацетіл-КоА, оксалоацетати, а-кетоглютарат, фумарат, сукцинат і кінцеві продукти азотистого обміну - сечовину, аміак, сечову к-ту і ін. В такому випадку під час гідролізу однієї молекули пірувату завдяки виділенню енергії відновлюється 18 молекули АТФ в циклі Кребса (цикл трикарбонових кислот).
АТФ як єдина й універсальна енергетична речовина. Усі прояви життєдіяльності, усі функції клітини потребують затрати енергії. Енергія потрібна для руху, для біосиптичних реакцій, перенесення речовин через клітинні мембрани, для будь-яких форм клітинної активності.
Джерелом енергії в живих клітинах, що забезпечує всі види їх діяльності, є аденозинтрифосфорна кислота (АТФ). Енергія, яка вивільняється під час розчеплення АТФ, забезпечує будь-які види клітинних функцій – рух, біосинтез, перенесення речовин через мембрани тощо. Оскільки запас АТФ у клітинні невеликий, то зрозуміло, що в міру зменшення кількості АТФ вміст її повинен відновлюватися. Насправді так і відбувається. Біологічний зміст решти реакцій енергетичного обміну в тому й полягає, що енергія, яка вивільняється внаслідок хімічних реакцій окислення вуглеводів та інших речовин, використовується для синтезу АТФ, тобто для поповнення її запису в клітині. Під час посиленої, але короткочасної роботи, наприклад під час бігу на коротку дистанцію, м’язи працюють майже виключно за рахунок розпаду АТФ, що міститься в них. Після закінчення бігу спортсмен посилено дихає, розігрівається: у цей період інтенсивно окислюються вуглеводи та інші речовини для поповнення витраченої АТФ. Від тривалої і не дуже напруженої роботи вміст АТФ у клітинах може істотно не змінюватися, оскільки реакції окислення встигають забезпечити швидке й повне відновлення втраченої АТФ.
Отже, АТФ – єдине й універсальне джерело енергії для функціональної діяльності клітини.
АТФ синтезується переважно в мітохондріях. Саме через це мітохондрії називають «силовими станціями» клітини. Утворена тут АТФ по каналах ендоплазматичної сітки надходить до тих ділянок клітини, де є потреба в енергії.
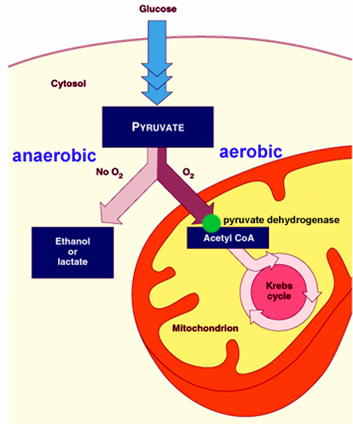
Етапи катаболізму або енергетичного обміну речовин.
1. Катаболізм у людей починається з ротової порожнини (амілаза розщеплює крохмаль).
2. Потім проходить розщеплення органічних сполук до мономерів у кишечнику (білки до амінокислот, ліпіди до жирних кислот, гліцерину й фосфорної кислоти, полісахариди до моносахаридів, нуклеїнові кислоти до нуклеотидів). Перший етап – підготовчий. На цьому етапі більші молекули вуглеводів, жирів, білків, нуклєїнових кислот розпадаються на невеликі молекули : з крохмалю утворюється глюкоза, з жирів – гліцерин і жирні кислоти, з білків – амінокисоти, з нуклеїнових кислот – нуклеотиди. Розпад речовин на цьому етапі супроводжується незначним енергетичним ефектом. Уся вивільнена при цьому енергія розсіються у вигляді тепла.
3. Далі після всмоктування сполук через кишечник вони потрапляють до клітин організму де і проходять розщеплювання до вищезазначених сполук (в гліколізі глюкоза розщеплюється до молочної або піровіноградної кислоти, у гідролізі піровіноградна кислота розщеплюється до СО2, Н2О та Н202 ). Другий етап енергетичного обміну називають безкисневим або неповим. Речовини, які утворилися на підготовчому етапі, – глюкоза, гліцерин, органічні кислоти, амінокислоти тощо – розпадаються далі. Це складний, багатоступінчастий процес. Він складається з ряду ферментативних реакцій, які йдуть одна за одною. Ферменти, що обслуговують цей процес, розміщені на внутрішньоклітинних мембранах правильними рядами. Речовина, яка надійшла на перший фермент цього ряду, переміщується, ніби на конвеєрі, на другий фермент, потім –третій і т.д. Завдяки цьому процес відбувається швидко й ефектвно. Розглянемо його на прикладі безкисневого розчеплення глюкози, яке називається гліколізом. Гліколіз – це ряд послідовних ферментативних реакцій. Його обслуговують 13 різних ферментів і в ході його утворюється більше 10 проміжних речовин. Багато проміжних реакцій гліколізу відбувається за участю фосфорної кислоти Н2РО4 і АДФ, а на кінці з нього сходять тривуглецева молочна кислота, АТФ і вода. Сумарне рівняння гліколізу записується так: С6Н12О6 + 2Н3РО4 + 2АДФ = 2С3Н6О3 + 2АТФ + 2Н2О.
Процес глікозу відбувається в усіх тваринних клітинах і в деяких мікроорганізмів. Усім відоме молочнокислотне бродіння (скисання молока, утворення сметани) спричиняється молочнокислими грибами і бактеріями. За механізмом воно цілком тотожне гліколізу. При цьому молочнокисле бродіння є подібним до спиртового бродіння. У обох випадках одним з продуктів гліколізу є спирт.
У рослинних клітин і в деяких дріжджових грибів глюкоза розпадається внаслідок спиртового бродіння. Спиртове бродіння, як і гліколіз, – довгий ряд ферментативних реакцій, причому більша частина реацій гліколізу й спиртового бродіння повністю збігаються, і тільки на останніх етапах є деякі відмінності. У ряді проміжних реакцій спиртового бродіння, як і в гліколізі, беруть участь Н3РО4 і АДФ. Кінцевими продуктами спиртового бродіння є оксид вуглецю (ІV), етиловий спирт, АТФ і вода. Сумарне рівняння спиртового бродіння записують так:
С6Н12О6 + 2Н3РО4 + 2АДФ = 2СО2 + 2С2Н5ОН + 2АТФ + 2Н2О.
З наведених рівнянь гліколізу й спиртового бродіння видно, що в цих процесах не бере участі кисень, тому їх і називають безкисневими або процесами з неповним розщепленням, оскільки повне розщеплення – це розщеплення до кінця, тобто перетворення глюкози в найпростіші сполуки – СО2 і Н2О, що відповідає рівнянню: С6Н12О6 + 6О2 = 6СО2 + 6Н2О.
Нарешті, і це особливо важливо, з рівнянь випливає, що від розпаду однієї молекули глюкози в ході гліколізу й спиртового бродіння утворюються дві молекули АТФ. Отже, розпад глюкози в процесі гліколізу й спиртового бродіння пов’язаний із синтезом універсальної енергетичної речовини АТФ.
Оскільки синтез АТФ – ендотермічний процес, то, очевидно, енергія для синтезу АТФ черпається з реакції безкисневого розщеплення глюкози. Отже, енергія, яка вивільняється в реакціях гліколізу, не вся переходить у тепло. Частина її використовується на синтез двох багатих на енергією фосфорних зв’язків.
Частково прості органічні сполуки не розщеплюються, а використовуються в анаболічних процесах.
В ході анаболічних перетворень відбувається біосинтез складних молекул і простих молекул-попередників. Автотрофні організми (зелені рослини і деякі бактерії) можуть здійснювати первинний синтез органіч. сполук з СО2 з використанням енергії сонячного світла (фотосинтез) або енергії окислення неорганічних речовин. Гетеротрофи синтезують органічні сполуки тільки за рахунок енергії і продуктів, що утворюються в результаті катаболічних перетворень. Початковою сировиною для процесів біосинтезу в цьому випадку служить невелике число сполук, в т.ч. ацетіл-КоА, сукциніл-КоА, рібоза, піровиноградна к-та, гліцерин, гліцин, аспарагінова, глютамінова і ін. амінокислоти. Кожна клітина синтезує характерні для неї білки, жири, вуглеводи і ін. сполуки. Напр. глікоген м'язів синтезується в м'язових клітинах, а не доставляється кров'ю з печінки. Як правило синтез включає декілька етапів і супроводжується споживанням енергії. Катаболізм і анаболізм протікають одночасно, стадія катаболіч. перетворень є початковою стадією анаболізму. Проте катаболіч. і анаболіч. шляхи О. р. не співпадають між собою. Напр., в розщеплюванні глікогену до молочної к-ти бере участь 12 ферментів, кожний з яких каталізує свій етап цього процесу. Синтез же глікогену з молочної к-ти включає тільки 9 ферментативних етапів, що відповідають етапам катаболізму, а 3 не дістають і замінюються іншими ферментативними реакціями, які використовуються тільки для біосинтезу. Не співпадають катаболіч. і анаболіч. шляхи обміну між білками і амінокислотами або між жирними к-тами і ацетіл-КоА. Більш того, різні. обмінні реакції приурочені певним ділянкам клітини. Вся ферментативна система гліколізу локалізується в розчинній фракції цитоплазми. У мітохондріях зосереджені процеси, пов'язані з біол. окисленням і окис. фосфорилуванням, в лізосомах- гидролітіч. ферменти. Процеси біосинтезу білка здійснюються в рібосомах, а біосинтезу ліпідів - в ендоплазматичній сітці і т.д. В різних частинах клітин локалізуються і хімічно несумісні реакції. Напр., окислення жирних к-т каталізується набором ферментів, локалізованих в мітохондріях, тоді як синтез жирних к-т з ацетіл-КоА - з допомогою іншого набору ферментів, локалізованих в цитоплазмі. Хоч і катаболіч., і анаболіч. шляхи здійснюються специфічними наборами ферментів, їх постійно зв'язують і загальні стадії О. р. Наїб, важливим загальним проміжним продуктом О. р., що бере участь у всіх процесах, є ацетіл-Ко А. Велике значення має цикл перетворень (цикл трикарбонових к-т), в ході якого ацетіл-КоА через ряд проміжних продуктів окиснюється повністю до CO2 і H2O. В той же час з ацетіл-Ко А починається синтез жирних к-т, холестерину, ряду азотовмісних сполук і т.д. В процесі еволюції організми виробили тонкі регуляторні системи що забезпечують високий ступінь впорядкованості і узгодженості реакцій і що дозволяють пристосуватися до змін умов навколишнього середовища. Для всіх організмів існують однакові системи регуляції, що діють на рівні клітинного О. р. В цьому випадку інтенсивність і спрямованість біохімічних реакцій може регулюватися дією або на активність ферменту шляхом його інгібування або активування, або на його синтез або деградацію. Велику роль в регуляції грає строга впорядкованість розташування ферментів в клітинних структурах, а також вибіркова проникність біол. мембран. Високорозвинені організми володіють додатковими регуляторними механізмами - нервовими і гормональними. Атрофія тканин після денервації указує на важливе значення нервових імпульсів для клітинного О. р. Гормони виконують в клітинах і тканинах контролюючі функції, або безпосередньо впливаючи на ферменти або їх синтез, або впливаючи на проникність клітинних мембран, функціональний стан клітинних органоїдів і систему цикліч. нуклеотідов.
4. Третій етап енергетичного обміну – стадія кисневого, або повного, розщеплення – дихання. Продукти, які виникли в попередній стадії, окислюються до кінця, тобто до СО2 і Н2О.
Основна умова здійснення цього процесу – наявність у навколишньому середовищі кисню і вбирання його клітиною. Стадія кисневого розщеплення, як і попередня стадія безкисневого розщеплення, являє собою ряд послідовних ферментативних реакцій. Кожна реакція каталізується особливим ферментом.
Увесь ферментативний ряд кисневого розщеплення зосереджений у мітохондріях, де ферменти розміщені на мембранах правильними рядами. Суть кожної з реакцій полягає в окислені органічної молекули, яка з кожним ступенем поступово руйнується і перетворюється на кінцеві продукти окислення: С2О і Н2О.
Усі проміжні реакції кисневого розщеплення, як і проміжні реакції безкисневого процесу, супроводжуються вивільненням енергії. Але кількість енергії, яка вивільняється на кожному ступені кисневого процесу, значно більша, ніж на кожному ступені безкисневого процесу. У сумі кисневого розщеплення дає величезну кількість енергії – 2600 кДж (650 ккал). Якби вся ця енергія вивільнилась в результаті однієї реакції, то клітина зазнала б теплового пошкодження. Оскільки процес поділяється на ряд проміжних ланок, такої загрози немає.
Докладне дослідження реакцій кисневого розщеплення показало, що в цих реакціях, як і в реакціях безкисневого процесу, беруть участь Н3РО4 і АДФ і що кисневий процес, як і безкисневий, нерозривно зв’язаний із синтезом АТФ. У процесі кисневого розщеплення двох тривуглецевих молекул утворюються 36 молекул АТФ – 36 багатих на енергію фосфатних зв’язків. Отже, сумарне рівняння кисневого процесу можна записати так:
2С3Н6О3 + 6О2 + 36Н3РО4 + 36АДФ = 6СО2 + 6Н2О + 36АТФ + 36Н2О,
а сумарне рівняння повного розщеплення глюкози так:
С6Н12О6 + 6О2 + 38Н3РО4 + 38АДФ = 6СО2 + 6Н2О + 38АТФ + 38Н2О.
Тепер стає зрозумілим значення для клітини третьої, кисневої стадії енергетичного обміну. Якщо в процесі безкисневого розщеплення глюкози вивільняються 200 кДж/моль (50 ккал/моль), то в стадії кисневого процесу вивільняються 2600 кДж (650 ккал), тобто в 13 разів більше. Якщо в процесі безкисневого розщеплення синтезуються дві молекули АТФ, то в кисневій стадії їх утворюється 36, тобто у 18 разів більше. Іншими словами, в наслідок розщеплення глюкози в клітині на стадії кисневого процесу вивільняється і перетворюється в інші форми енергії понад 90% енергії глюкози (Цикл Кребса).
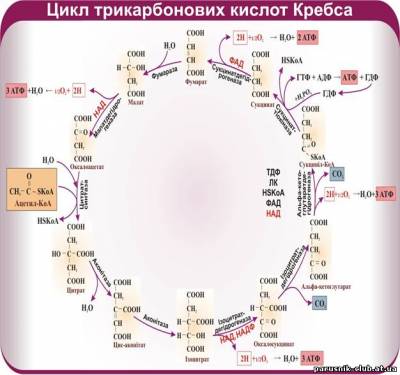
Зробимо знову розрахунки. Усього в процесі розщеплення глюкози до СО2 і Н2О, тобто в кисневому і безкисневому процесах, синтезуються 2 + 36 = 38 молекул АТФ. Таким чином, у потенціальну енергію АТФ переходить 38×40=1520 кДж (38×10=380 ккал). Усього від розщеплення глюкози ( в безкисневій і кисневій стадіях ) вивільняється 200+2600=2800 кДж (50+650=700 ккал). Отже майже 55% усієї енергії, яка вивільняється від розщеплення глюкози, зберігається клітиною у формі АТФ. Решта її (45%) розсіюється у вигляді тепла. Щоб оцінити значення цих цифр, нагадаємо, що в парових машинах з енергії, яка вивільняється внаслідок згоряння вугілля, у корисну форму перетворюється не більше як 12-15% . У двигунах внутрішнього згоряння коефіцієнт корисної дії досягає приблизно 35% . Таким чином, за ефективністю перетворення енергії жива клітина переважає всі відомі в техніці перетворювачі енергії.
Порівнявши кількість енергії, яка вивільняється в процесі безкисневого і кисневого розщеплення глюкози, а також кількості молекул АТФ, що синтезуються в обох стадіях, побачимо, що кисневий процес незрівнянно ефективніший, ніж безкисневий. У стадії безкисневого рощеплення вивільняється приблизно ¹/20 енергії, що звільняється в кисневому процесі. Цілком зрозуміло, що в нормальних умовах для мобілізації енергії в клітині завжди використовуються і безкисневий, і кисневий способи розщеплення. Якщо кисневий процес утруднений або зовсім неможливий, наприклад, коли не вистачає кисню, тоді для підтримання життя залишається тільки безкисневий процес. Але при цьому для добування АТФ у кількості, потрібній для життєдіяльності, клітині доводиться розщеплювати дуже багато глюкози.
МЕТАБОЛІЗМ (від греч. metabole- зміна, перетворення), 1) те ж, що обмін речовин. 2) У більш вузькому сенсі М.- проміжний обмін, що охоплює всю сукупність реакцій, головним чином ферментативних, які протікають в клітинах і забезпечують як розщеплювання складних сполук, так і їх синтез і взаємоперетворення.
Асиміляція – процеси, пов’язані з поглинанням із довкілля, засвоєнням і накопиченням хімічних речовин, які використовуються для синтезу сполук, подібних для організму, називають асиміляцією.
Енергетичний обмін – сукупність реакцій розщеплення складних сполук, які супроводжуються виділенням енергії.
Пластичний обмін – сукупність реакцій синтезу, що забезпечують ріст клітин і поновлення їхнього хімічного складу
Дисиміляція - процеси обміну речовин, які приводять до розкладу певних сполук в клітинах
Сукупність реакцій розчеплення називають енергетичним обміном клітин, або дисиміляцією. Дисиміляція прямо протилежна асиміляції: в результаті розщеплення речовини втрачають подібність до речовини клітини.
Пластичний і енергетичний обміни ( асиміляція і дисиміляція ) нерозривно зв’язані між собою. З одного боку, реакції біосинтезу потребують затрати енергії, яка черпається з реакцій розщеплення. З другого боку, для здійснення реакцій енергетичного обміну потрібний постійний біосинтез ферментів, що обслуговують ці реакції, бо в процесі своєї діяльності вони зношуються і руйнуються.
АВТОТРОФНІ ОРГАНІЗМИ, автотрофи (від авто... і ...троф), організми, що використовують для побудови свого тіла CO2 як єдине або головне. джерела вуглецю і що володіють як системою ферментів для асиміляції СОз, так і здатністю синтезувати всі компоненти клітини. Деякі А. о. можуть потребувати екзогенних (що поступають ззовні) вітамінів або чинників зростання. А. о. протиставляються ГЕТЕРОТРОФНИМ ОРГАНІЗМАМ що використовують екзогенні органічні речовини.
До А. о. відносяться:
1. наземні зелені рослини,
2. водорості,
3. фототрофні бактерії, здібні до фотосинтезу,
4. деякі бактерії що використовують окислення неорганічних речовин - хемоавтотрофи (див. Хемосинтез).
Більшість А. о. асимілює CO2 через відновний пентозофосфатний шлях. У деяких бактерій, наприклад, метанобразуючих, CO2 асимілюється іншим шляхом. Обговорюється можливість віднесення до А. о. бактерій, що використовують як джерело вуглецю метан. А. о.- первинні продуценти органічних речовини в біосфері, створюючі перший трофічний рівень у співтовариствах. Роль фотосинтезуючих А. о. в природі є таким, що визначає, оскільки вони утворюють основну масу органічної речовини в біосфері - быля. 162 • 109 т/год, в т.ч. 2/3 дають наземні рослини. Внесок інших груп А. о. кількісно невеликий. Діяльністю А. о. визначається як існування всіх інших організмів, так і хід біогеохімічних циклів в круговороті речовин в природі.
ОБМІН РЕЧОВИН, метаболізм, сукупність тих, що протікають в живих організмах хімічних перетворень що забезпечують їх зростання, життєдіяльність, відтворення, постійний контакт і обмін з навколишнім середовищем. Завдяки О. в. відбувається розщеплювання і синтез молекул, що входять в склад клітин, утворення, руйнування і оновлення клітинних структур і міжклітинної речовини. Наприклад у людини половина всіх тканинних білків розщеплюється і будується наново в середньому протягом 80 діб, білки печінки до сироватки крові наполовину оновлюються кожні 10 діб, а білки м’язів- 180, ферменты печінки - кожні 2-4 години. О. р. не можливо відокремити від процесів перетворення енергії: потенційна енергія хімічних зв'язків складних органічних молекул в результаті хімічних перетворень переходить в інші види енергії, використовуваної на синтез нових сполук, для підтримки структури і функції клітин, температури тіла, для здійснення роботи і т.д. Всі реакції О. в. і перетворення енергії протікають за участю біологічних каталізаторів - ферментів. У самих різних організмів О. в. відрізняється впорядкованістю і схожістю послідовності ферментативних перетворень не дивлячись на великий асортимент хімічних сполук, що залучаються до обміну. В той же час для кожного виду характерний особливий, генетично закріплений тип О. в., обумовлений умовами його існування. О. в. складається з двох взаємопов'язаних, що одночасно протікають в організмі процесів - асиміляції і дисиміляції, або анаболізму і катаболізму. В ході катаболічних перетворень відбувається розщеплювання великих органічних молекул до простих сполук з одночасним виділенням енергії, яка запасається у формі багатих енергією фосфатних зв'язків, головним чином. у молекулі аденозінтрифосфорної к-ти (АТФ) і інших багатих енергією з'єднань.
Катаболічні перетворення звичайно здійснюються в результаті гідролітичних і окиснювальних реакцій і протікають як у відсутність кисню (анаеробний шлях - гліколіз, бродіння), так і при його участі (аеробний шлях - дихання). В першому випадку під час гліколізу завдяки виділенню енергії відновлюється 2 молекули АТФ при рощепленні однієї молекули глюкози. Другий шлях еволюційно молодший і в енергетичному відношенні більш вигідний. Він забезпечує повне розщеплювання органічних молекул до CO2 і Н2О. Різноманітні органічні сполуки в ході катаболічних процесів перетворюються на обмежене число невеликих молекул (крім СО2 і Н2О): вуглеводи - в тріозофосфати і (або) пірувати (піровіноградна кислота), жири - в ацетіл-КоА пропіоніл-КоА і гліцерин, білки - в ацетіл-КоА, оксалоацетати, а-кетоглютарат, фумарат, сукцинат і кінцеві продукти азотистого обміну - сечовину, аміак, сечову к-ту і ін. В такому випадку під час гідролізу однієї молекули пірувату завдяки виділенню енергії відновлюється 18 молекули АТФ в циклі Кребса (цикл трикарбонових кислот).
АТФ як єдина й універсальна енергетична речовина. Усі прояви життєдіяльності, усі функції клітини потребують затрати енергії. Енергія потрібна для руху, для біосиптичних реакцій, перенесення речовин через клітинні мембрани, для будь-яких форм клітинної активності.
Джерелом енергії в живих клітинах, що забезпечує всі види їх діяльності, є аденозинтрифосфорна кислота (АТФ). Енергія, яка вивільняється під час розчеплення АТФ, забезпечує будь-які види клітинних функцій – рух, біосинтез, перенесення речовин через мембрани тощо. Оскільки запас АТФ у клітинні невеликий, то зрозуміло, що в міру зменшення кількості АТФ вміст її повинен відновлюватися. Насправді так і відбувається. Біологічний зміст решти реакцій енергетичного обміну в тому й полягає, що енергія, яка вивільняється внаслідок хімічних реакцій окислення вуглеводів та інших речовин, використовується для синтезу АТФ, тобто для поповнення її запису в клітині. Під час посиленої, але короткочасної роботи, наприклад під час бігу на коротку дистанцію, м’язи працюють майже виключно за рахунок розпаду АТФ, що міститься в них. Після закінчення бігу спортсмен посилено дихає, розігрівається: у цей період інтенсивно окислюються вуглеводи та інші речовини для поповнення витраченої АТФ. Від тривалої і не дуже напруженої роботи вміст АТФ у клітинах може істотно не змінюватися, оскільки реакції окислення встигають забезпечити швидке й повне відновлення втраченої АТФ.
Отже, АТФ – єдине й універсальне джерело енергії для функціональної діяльності клітини.
АТФ синтезується переважно в мітохондріях. Саме через це мітохондрії називають «силовими станціями» клітини. Утворена тут АТФ по каналах ендоплазматичної сітки надходить до тих ділянок клітини, де є потреба в енергії.
1. Катаболізм у людей починається з ротової порожнини (амілаза розщеплює крохмаль).
2. Потім проходить розщеплення органічних сполук до мономерів у кишечнику (білки до амінокислот, ліпіди до жирних кислот, гліцерину й фосфорної кислоти, полісахариди до моносахаридів, нуклеїнові кислоти до нуклеотидів). Перший етап – підготовчий. На цьому етапі більші молекули вуглеводів, жирів, білків, нуклєїнових кислот розпадаються на невеликі молекули : з крохмалю утворюється глюкоза, з жирів – гліцерин і жирні кислоти, з білків – амінокисоти, з нуклеїнових кислот – нуклеотиди. Розпад речовин на цьому етапі супроводжується незначним енергетичним ефектом. Уся вивільнена при цьому енергія розсіються у вигляді тепла.
3. Далі після всмоктування сполук через кишечник вони потрапляють до клітин організму де і проходять розщеплювання до вищезазначених сполук (в гліколізі глюкоза розщеплюється до молочної або піровіноградної кислоти, у гідролізі піровіноградна кислота розщеплюється до СО2, Н2О та Н202 ). Другий етап енергетичного обміну називають безкисневим або неповим. Речовини, які утворилися на підготовчому етапі, – глюкоза, гліцерин, органічні кислоти, амінокислоти тощо – розпадаються далі. Це складний, багатоступінчастий процес. Він складається з ряду ферментативних реакцій, які йдуть одна за одною. Ферменти, що обслуговують цей процес, розміщені на внутрішньоклітинних мембранах правильними рядами. Речовина, яка надійшла на перший фермент цього ряду, переміщується, ніби на конвеєрі, на другий фермент, потім –третій і т.д. Завдяки цьому процес відбувається швидко й ефектвно. Розглянемо його на прикладі безкисневого розчеплення глюкози, яке називається гліколізом. Гліколіз – це ряд послідовних ферментативних реакцій. Його обслуговують 13 різних ферментів і в ході його утворюється більше 10 проміжних речовин. Багато проміжних реакцій гліколізу відбувається за участю фосфорної кислоти Н2РО4 і АДФ, а на кінці з нього сходять тривуглецева молочна кислота, АТФ і вода. Сумарне рівняння гліколізу записується так: С6Н12О6 + 2Н3РО4 + 2АДФ = 2С3Н6О3 + 2АТФ + 2Н2О.
Процес глікозу відбувається в усіх тваринних клітинах і в деяких мікроорганізмів. Усім відоме молочнокислотне бродіння (скисання молока, утворення сметани) спричиняється молочнокислими грибами і бактеріями. За механізмом воно цілком тотожне гліколізу. При цьому молочнокисле бродіння є подібним до спиртового бродіння. У обох випадках одним з продуктів гліколізу є спирт.
У рослинних клітин і в деяких дріжджових грибів глюкоза розпадається внаслідок спиртового бродіння. Спиртове бродіння, як і гліколіз, – довгий ряд ферментативних реакцій, причому більша частина реацій гліколізу й спиртового бродіння повністю збігаються, і тільки на останніх етапах є деякі відмінності. У ряді проміжних реакцій спиртового бродіння, як і в гліколізі, беруть участь Н3РО4 і АДФ. Кінцевими продуктами спиртового бродіння є оксид вуглецю (ІV), етиловий спирт, АТФ і вода. Сумарне рівняння спиртового бродіння записують так:
С6Н12О6 + 2Н3РО4 + 2АДФ = 2СО2 + 2С2Н5ОН + 2АТФ + 2Н2О.
З наведених рівнянь гліколізу й спиртового бродіння видно, що в цих процесах не бере участі кисень, тому їх і називають безкисневими або процесами з неповним розщепленням, оскільки повне розщеплення – це розщеплення до кінця, тобто перетворення глюкози в найпростіші сполуки – СО2 і Н2О, що відповідає рівнянню: С6Н12О6 + 6О2 = 6СО2 + 6Н2О.
Нарешті, і це особливо важливо, з рівнянь випливає, що від розпаду однієї молекули глюкози в ході гліколізу й спиртового бродіння утворюються дві молекули АТФ. Отже, розпад глюкози в процесі гліколізу й спиртового бродіння пов’язаний із синтезом універсальної енергетичної речовини АТФ.
Оскільки синтез АТФ – ендотермічний процес, то, очевидно, енергія для синтезу АТФ черпається з реакції безкисневого розщеплення глюкози. Отже, енергія, яка вивільняється в реакціях гліколізу, не вся переходить у тепло. Частина її використовується на синтез двох багатих на енергією фосфорних зв’язків.
Частково прості органічні сполуки не розщеплюються, а використовуються в анаболічних процесах.
В ході анаболічних перетворень відбувається біосинтез складних молекул і простих молекул-попередників. Автотрофні організми (зелені рослини і деякі бактерії) можуть здійснювати первинний синтез органіч. сполук з СО2 з використанням енергії сонячного світла (фотосинтез) або енергії окислення неорганічних речовин. Гетеротрофи синтезують органічні сполуки тільки за рахунок енергії і продуктів, що утворюються в результаті катаболічних перетворень. Початковою сировиною для процесів біосинтезу в цьому випадку служить невелике число сполук, в т.ч. ацетіл-КоА, сукциніл-КоА, рібоза, піровиноградна к-та, гліцерин, гліцин, аспарагінова, глютамінова і ін. амінокислоти. Кожна клітина синтезує характерні для неї білки, жири, вуглеводи і ін. сполуки. Напр. глікоген м'язів синтезується в м'язових клітинах, а не доставляється кров'ю з печінки. Як правило синтез включає декілька етапів і супроводжується споживанням енергії. Катаболізм і анаболізм протікають одночасно, стадія катаболіч. перетворень є початковою стадією анаболізму. Проте катаболіч. і анаболіч. шляхи О. р. не співпадають між собою. Напр., в розщеплюванні глікогену до молочної к-ти бере участь 12 ферментів, кожний з яких каталізує свій етап цього процесу. Синтез же глікогену з молочної к-ти включає тільки 9 ферментативних етапів, що відповідають етапам катаболізму, а 3 не дістають і замінюються іншими ферментативними реакціями, які використовуються тільки для біосинтезу. Не співпадають катаболіч. і анаболіч. шляхи обміну між білками і амінокислотами або між жирними к-тами і ацетіл-КоА. Більш того, різні. обмінні реакції приурочені певним ділянкам клітини. Вся ферментативна система гліколізу локалізується в розчинній фракції цитоплазми. У мітохондріях зосереджені процеси, пов'язані з біол. окисленням і окис. фосфорилуванням, в лізосомах- гидролітіч. ферменти. Процеси біосинтезу білка здійснюються в рібосомах, а біосинтезу ліпідів - в ендоплазматичній сітці і т.д. В різних частинах клітин локалізуються і хімічно несумісні реакції. Напр., окислення жирних к-т каталізується набором ферментів, локалізованих в мітохондріях, тоді як синтез жирних к-т з ацетіл-КоА - з допомогою іншого набору ферментів, локалізованих в цитоплазмі. Хоч і катаболіч., і анаболіч. шляхи здійснюються специфічними наборами ферментів, їх постійно зв'язують і загальні стадії О. р. Наїб, важливим загальним проміжним продуктом О. р., що бере участь у всіх процесах, є ацетіл-Ко А. Велике значення має цикл перетворень (цикл трикарбонових к-т), в ході якого ацетіл-КоА через ряд проміжних продуктів окиснюється повністю до CO2 і H2O. В той же час з ацетіл-Ко А починається синтез жирних к-т, холестерину, ряду азотовмісних сполук і т.д. В процесі еволюції організми виробили тонкі регуляторні системи що забезпечують високий ступінь впорядкованості і узгодженості реакцій і що дозволяють пристосуватися до змін умов навколишнього середовища. Для всіх організмів існують однакові системи регуляції, що діють на рівні клітинного О. р. В цьому випадку інтенсивність і спрямованість біохімічних реакцій може регулюватися дією або на активність ферменту шляхом його інгібування або активування, або на його синтез або деградацію. Велику роль в регуляції грає строга впорядкованість розташування ферментів в клітинних структурах, а також вибіркова проникність біол. мембран. Високорозвинені організми володіють додатковими регуляторними механізмами - нервовими і гормональними. Атрофія тканин після денервації указує на важливе значення нервових імпульсів для клітинного О. р. Гормони виконують в клітинах і тканинах контролюючі функції, або безпосередньо впливаючи на ферменти або їх синтез, або впливаючи на проникність клітинних мембран, функціональний стан клітинних органоїдів і систему цикліч. нуклеотідов.
4. Третій етап енергетичного обміну – стадія кисневого, або повного, розщеплення – дихання. Продукти, які виникли в попередній стадії, окислюються до кінця, тобто до СО2 і Н2О.
Основна умова здійснення цього процесу – наявність у навколишньому середовищі кисню і вбирання його клітиною. Стадія кисневого розщеплення, як і попередня стадія безкисневого розщеплення, являє собою ряд послідовних ферментативних реакцій. Кожна реакція каталізується особливим ферментом.
Увесь ферментативний ряд кисневого розщеплення зосереджений у мітохондріях, де ферменти розміщені на мембранах правильними рядами. Суть кожної з реакцій полягає в окислені органічної молекули, яка з кожним ступенем поступово руйнується і перетворюється на кінцеві продукти окислення: С2О і Н2О.
Усі проміжні реакції кисневого розщеплення, як і проміжні реакції безкисневого процесу, супроводжуються вивільненням енергії. Але кількість енергії, яка вивільняється на кожному ступені кисневого процесу, значно більша, ніж на кожному ступені безкисневого процесу. У сумі кисневого розщеплення дає величезну кількість енергії – 2600 кДж (650 ккал). Якби вся ця енергія вивільнилась в результаті однієї реакції, то клітина зазнала б теплового пошкодження. Оскільки процес поділяється на ряд проміжних ланок, такої загрози немає.
Докладне дослідження реакцій кисневого розщеплення показало, що в цих реакціях, як і в реакціях безкисневого процесу, беруть участь Н3РО4 і АДФ і що кисневий процес, як і безкисневий, нерозривно зв’язаний із синтезом АТФ. У процесі кисневого розщеплення двох тривуглецевих молекул утворюються 36 молекул АТФ – 36 багатих на енергію фосфатних зв’язків. Отже, сумарне рівняння кисневого процесу можна записати так:
2С3Н6О3 + 6О2 + 36Н3РО4 + 36АДФ = 6СО2 + 6Н2О + 36АТФ + 36Н2О,
а сумарне рівняння повного розщеплення глюкози так:
С6Н12О6 + 6О2 + 38Н3РО4 + 38АДФ = 6СО2 + 6Н2О + 38АТФ + 38Н2О.
Тепер стає зрозумілим значення для клітини третьої, кисневої стадії енергетичного обміну. Якщо в процесі безкисневого розщеплення глюкози вивільняються 200 кДж/моль (50 ккал/моль), то в стадії кисневого процесу вивільняються 2600 кДж (650 ккал), тобто в 13 разів більше. Якщо в процесі безкисневого розщеплення синтезуються дві молекули АТФ, то в кисневій стадії їх утворюється 36, тобто у 18 разів більше. Іншими словами, в наслідок розщеплення глюкози в клітині на стадії кисневого процесу вивільняється і перетворюється в інші форми енергії понад 90% енергії глюкози (Цикл Кребса).
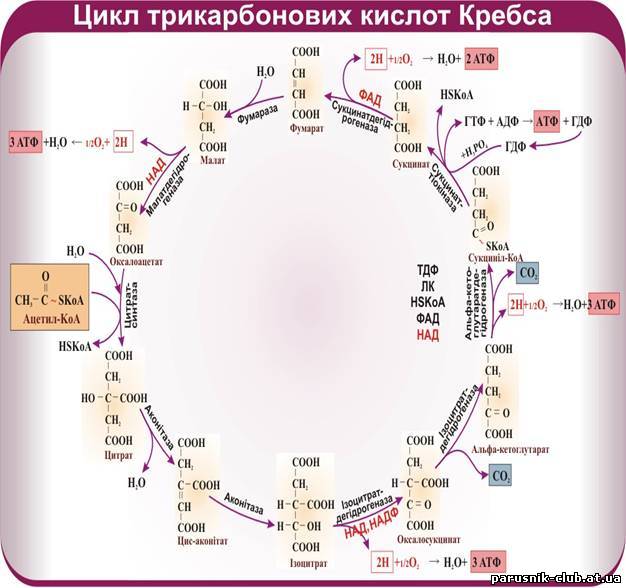
Порівнявши кількість енергії, яка вивільняється в процесі безкисневого і кисневого розщеплення глюкози, а також кількості молекул АТФ, що синтезуються в обох стадіях, побачимо, що кисневий процес незрівнянно ефективніший, ніж безкисневий. У стадії безкисневого рощеплення вивільняється приблизно ¹/20 енергії, що звільняється в кисневому процесі. Цілком зрозуміло, що в нормальних умовах для мобілізації енергії в клітині завжди використовуються і безкисневий, і кисневий способи розщеплення. Якщо кисневий процес утруднений або зовсім неможливий, наприклад, коли не вистачає кисню, тоді для підтримання життя залишається тільки безкисневий процес. Але при цьому для добування АТФ у кількості, потрібній для життєдіяльності, клітині доводиться розщеплювати дуже багато глюкози.






