

Лекція № 6: Нуклеїнові кислоти.
«Это долгая история,
которой не следовало
бы быть долгой,
но понадобилось бы
слишком много времени,
чтобы сделать ее короткой.
Генри Дэвид Торо
І. Будова нуклеїнових кислот, які несуть генетичну інформацію.
ІІ. Функції нуклеїнових кислот, які несуть генетичну інформацію.
ІІІ. Будова нуклеїнових кислот, які не несуть генетичну інформацію.
ІV. Функції нуклеїнових кислот, які не несуть генетичну інформацію.
І. Будова нуклеїнових кислот, які несуть генетичну інформацію.
ДЕЗОКСИРИБОНУКЛЕЇНОВІ КИСЛОТИ (ДНК), нуклеїнові к-ти, що містять в якості вуглеводного компоненту дезоксирибозу, а як азотисті основи аденін (А), гуанін (Г), цитозін (Ц), тимін (Т).
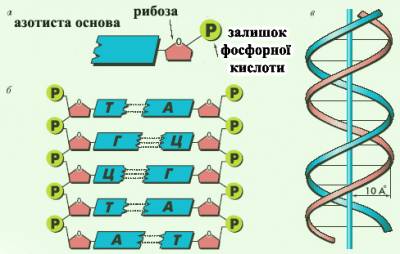
Рис. 1. Будова фрагменту ДНК (декілька нуклеотидів).
Присутні в клітинах будь-якого організму, а також входять до складу вірусів. Первинна структура молекули ДНК (подібно до первинної структури білка) індивідуальна і специфічна для кожної природної ДНК і представляє кодову форму запису біологічної інформації (генетичний код). Вперше доказ генетичної ролі ДНК одержано в 1944 О. Эйвері із співробітниками (США) в дослідах по трансформації здійснених на бактеріях. У вигляді унікальної послідовності нуклеотидів (які містять один залишок азотистої основи, дезоксирибозу та один залишок фосфорної кислоти) інформація про структуру білка зберігається в частинах молекули ДНК і багато разів і точно відтворюється за допомогою механізмів реплікації і транскрипції, потім в процесі синтезу білків на рібосомах (трансляція) реалізується в послідовність амінокислот. Нуклеотідний склад ДНК, виділеної з організмів різних видів, сильно розрізняється, але є характерним для кожного виду. Відоспецифічність ДНК - основа геносистематіки і використовується для встановлення філогенетичної близькості організмів. Вміст нуклеотідов в ДНК підкоряється закономірностям, розкритим Э. Чаргаффом (1950): сумарна кількість пурінових основ рівна сумі пірімідінових основ, причому кількість А рівне кількості Т, а кількість Г- кількості Ц. Ці закономірності визначаються особливостями макромолекул структури ДНК, відкритої Дж. Уотсоном і Ф. Кріком (1953). Згідно розробленої ними тривимірної моделі структури ДНК молекули ДНК представляють дві правозакручені навколо загальної осі спіральні полінуклеотидні ланцюги з кроком спіралі 34 А, що містять 10 нуклеотидів на віток і розташовані антіпаралельно (послідовність міжнуклеотидних зв'язків в двох ланцюгах направлена в протилежні боки) на відстані 18 А один від одного. Фосфатні групи знаходяться на зовнішніх. сторонах подвійної спіралі, а азотисті основи - усередині, таким чином, що їх площини перпендикулярні осі молекули. Основи, що при цьому протилежать, в ланцюгах утворюють за рахунок водневих зв'язків т.з. комплементарні пари А-Т і Г-Ц. Т. ч., послідовність основ в одному ланцюгу однозначно визначає послідовність основ в іншому комплементарному ланцюзі молекули.
Більшість природної ДНК має дволанцюгову структуру, лінійну або кільцеву форму (у останньому випадку кінці молекули ковалентно замкнуті). Виняток становлять деякі віруси, в складі яких виявлена одноланцюгова ДНК, також лінійна або кільцева. Біспіральна структура не є абсолютно жорсткою, що робить можливим утворення перегинів, петель, суперспіралей, необхідних для пакування гігантських молекул ДНК в малому об'ємі клітини або вірусу. У клітинах прокаріот ДНК організована в одну хромосому - нуклеоїд і представляє єдину макромолекулу, упаковану у вигляді суперспіралізованих петель; невеликі циклічні молекули ДНК присутні в плазмідах – шматочках ДНК, які не входять у структуру основної циклічної ДНК. ДНК (кільцеві молекули з мол. м. 106-107) входять до складу мітохондрій н хлоропластів, де забезпечують автономний синтез білків в цих клітинних органоїдах. Мінімальна для даного виду кількість ДНК містять статеві клітки, що мають галоїдний набір хромосом. У ядрах соматичних клітин ДНК, як правило, удвічі більше, ніж відповідає диплоїдному набору. Окрім водневих зв'язків стабілізація спіральної структури ДНК досягається також міжплощинними взаємодіями основ. Параметри моделі Уотсона - Крика відповідають конформації ДНК у фізіологічних умовах (т.з. В-форма ДНК). Нагрівання, зміна рН, пониження іонної сили, ряд інших чинників викликають денатурацію дволанцюгової молекули ДНК.
Процес реплікації (самоподвоєння) ДНК
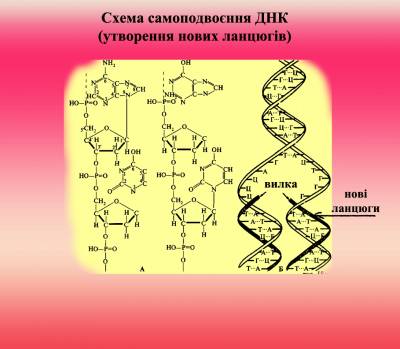
Рис. 2. Процес реплікаціх ДНК
відбувається за допомогою ферментів геліказ, які розривають водневі зв’язки між ланцюгами та ДНК-полімераз, які утворюють комплементарний новий ланцюг на матриці (одному з існуючих ланцюгів). Реплікація хромосомної ДНК в клітині, що ділиться, починається з локального розплітання подвійної спіралі і утворення реплікативної вилки, в чому беруть участь специфічні ендонуклеази що розплітають білки. Синхронність реплікації обох антипаралельних ланцюгів забезпечується завдяки тому, що синтез йде короткими фрагментами (100-10 000 нуклеотідов), які приєднуються потім до ланцюгів. А. Корнберг в 1967 здійснив ферментативний синтез біол. активної ДНК in vitro.
Процес транскрипції – зчитування інформації з ДНК на РНК (рибонуклеїнову кислоту) відбувається також за допомогою геліказ і РНК-полімераз. При цьому формуються інформаційна РНК (і - РНК) – яка несе інформацію про відповідну послідовність амінокислот в протеїнах, складається з 300 – 30000 нуклеотидів, транспортна РНК (т-РНК) – яка несе інформацію за допомогою триплету (трьох нуклеотидів) про відповідну амінокислоту, складається з 70 – 90 нуклеотидів, рибосомна РНК – яка забезпечує певне просторове розташування іРНК і тРНК на рибосомі, становить близько 85 % загальної кількості РНК клітини і 60 % маси рибосоми.
ІІ. Функції нуклеїнових кислот, які несуть генетичну інформацію.
Роль ДНК полягає в збереженні інформації про будову організму (в першу чергу білків, їх співвідношення та розташування в організмі). Ця інформація специфічна для кожного біологічного виду.
Роль і.РНК – це перенос генетичної інформації від зони її збереження (тобто від ДНК) до міста перетворення інформації в біомолекули (білки). В процесі перетворення інформації в біомолекули приймають участь також т. РНК – молекула, яка підносить до місць синтезу білків амінокислоти і р.РНК – яка складає приблизно 60 % рибосоми – органели, на якій відбувається біосинтез білків.
Таким чином існують три види нуклеїнових кислот:
1. і. РНК - інформаційна;
2. т. РНК - транспортна;
3. р. РНК - рибосомна.
ІІІ. Будова нуклеїнових кислот, які не несуть генетичну інформацію.
ІV. Функції нуклеїнових кислот, які не несуть генетичну інформацію.
АТФ - нуклеїнові кислоти, які не несуть генетичної інформації, але виконують функцію накопичення енергії під час утворення хімічних зв’язків своїх молекул. АТФ – аденозінтрифосфорна кислота, АДФ – аденозіндифосфорна кислота, АМФ – аденозінмонофосфорна кислота (рис. 3).
АТФ, АДФ, АМФ – нуклеотиди, моно, ди, або три фосфорні ефіри аденіну. Мають в складі аденін, рибозу, один або два, або три залишки фосфорної кислоти.
АТФ ресинтезуються у еукаріотів переважно у мітохондріях, відновлюючись з АМФ та АДФ. У прокаріотів відновлення АТФ відбувається переважно у клітинній стінці.
НАД (НІКОТИНАМІДАДЕНІНДИНУКЛЕОТИД) динуклеотид, що складається з аденина, аміда нікотинової кислоти, двох залишків рибози і двох залишків фосфорної кислоти; кофермент дегідрогенази, виявлений у всіх живих клітинах, що функціонує на початкових етапах биологічного окислення жирів, білків і вуглеводів. Відкритий в 1904 в дріжджовому соку А. Гарденом і У. Йонгом; будова установлена в 1936 О. Варбургом і X. Эйлером-Хельпіном. С дегідрогеназами у реакціях, що каталізують, НАД і його фосфоріловане похідне НАДФ - проміжні акцептори і переносники електронів і водню. У клітинах НАД присутній в значно більших кількостях, чим НАДФ. У більшості тканин біосинтез НАД здійснюється багатоферментною системою як з нікотином, так і з нікотинової кіслоти; у печінці і нирках містяться ферментні системи, здатні синтезувати НАД з триптофану. НАДФ відрізняється від НАД наявністю третього залишку фосфорної кислоти. У клітинах НАДФ присутній в основному у відновленій формі (НАДФ.Н). Окислений НАДФ+ - акцептор водню при окисленні глюкозо-6-фосфату в пентозному циклі, в світлових реакціях фотосинтезу і т. і. Відновлений НАДФ Н використовується головним чином у біосинтезах жирних кислот, вуглеводів (у темнових реакціях фотосинтезу), відновному амінуванні альфа-кетоглутарової кислоти. При дії специфичної НАДФ-цитохромредуктази відбувається пряме окислення НАДФ у дыхальному ланцюзі. Біосинтез НАДФ здійснюється при фосфори-луванні НАД ферментом НАД-кіназою.
«Это долгая история,
которой не следовало
бы быть долгой,
но понадобилось бы
слишком много времени,
чтобы сделать ее короткой.
Генри Дэвид Торо
І. Будова нуклеїнових кислот, які несуть генетичну інформацію.
ІІ. Функції нуклеїнових кислот, які несуть генетичну інформацію.
ІІІ. Будова нуклеїнових кислот, які не несуть генетичну інформацію.
ІV. Функції нуклеїнових кислот, які не несуть генетичну інформацію.
І. Будова нуклеїнових кислот, які несуть генетичну інформацію.
ДЕЗОКСИРИБОНУКЛЕЇНОВІ КИСЛОТИ (ДНК), нуклеїнові к-ти, що містять в якості вуглеводного компоненту дезоксирибозу, а як азотисті основи аденін (А), гуанін (Г), цитозін (Ц), тимін (Т).
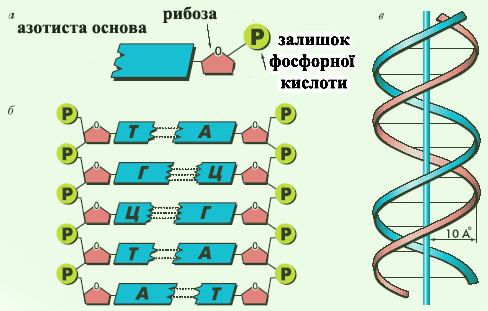
Присутні в клітинах будь-якого організму, а також входять до складу вірусів. Первинна структура молекули ДНК (подібно до первинної структури білка) індивідуальна і специфічна для кожної природної ДНК і представляє кодову форму запису біологічної інформації (генетичний код). Вперше доказ генетичної ролі ДНК одержано в 1944 О. Эйвері із співробітниками (США) в дослідах по трансформації здійснених на бактеріях. У вигляді унікальної послідовності нуклеотидів (які містять один залишок азотистої основи, дезоксирибозу та один залишок фосфорної кислоти) інформація про структуру білка зберігається в частинах молекули ДНК і багато разів і точно відтворюється за допомогою механізмів реплікації і транскрипції, потім в процесі синтезу білків на рібосомах (трансляція) реалізується в послідовність амінокислот. Нуклеотідний склад ДНК, виділеної з організмів різних видів, сильно розрізняється, але є характерним для кожного виду. Відоспецифічність ДНК - основа геносистематіки і використовується для встановлення філогенетичної близькості організмів. Вміст нуклеотідов в ДНК підкоряється закономірностям, розкритим Э. Чаргаффом (1950): сумарна кількість пурінових основ рівна сумі пірімідінових основ, причому кількість А рівне кількості Т, а кількість Г- кількості Ц. Ці закономірності визначаються особливостями макромолекул структури ДНК, відкритої Дж. Уотсоном і Ф. Кріком (1953). Згідно розробленої ними тривимірної моделі структури ДНК молекули ДНК представляють дві правозакручені навколо загальної осі спіральні полінуклеотидні ланцюги з кроком спіралі 34 А, що містять 10 нуклеотидів на віток і розташовані антіпаралельно (послідовність міжнуклеотидних зв'язків в двох ланцюгах направлена в протилежні боки) на відстані 18 А один від одного. Фосфатні групи знаходяться на зовнішніх. сторонах подвійної спіралі, а азотисті основи - усередині, таким чином, що їх площини перпендикулярні осі молекули. Основи, що при цьому протилежать, в ланцюгах утворюють за рахунок водневих зв'язків т.з. комплементарні пари А-Т і Г-Ц. Т. ч., послідовність основ в одному ланцюгу однозначно визначає послідовність основ в іншому комплементарному ланцюзі молекули.
Більшість природної ДНК має дволанцюгову структуру, лінійну або кільцеву форму (у останньому випадку кінці молекули ковалентно замкнуті). Виняток становлять деякі віруси, в складі яких виявлена одноланцюгова ДНК, також лінійна або кільцева. Біспіральна структура не є абсолютно жорсткою, що робить можливим утворення перегинів, петель, суперспіралей, необхідних для пакування гігантських молекул ДНК в малому об'ємі клітини або вірусу. У клітинах прокаріот ДНК організована в одну хромосому - нуклеоїд і представляє єдину макромолекулу, упаковану у вигляді суперспіралізованих петель; невеликі циклічні молекули ДНК присутні в плазмідах – шматочках ДНК, які не входять у структуру основної циклічної ДНК. ДНК (кільцеві молекули з мол. м. 106-107) входять до складу мітохондрій н хлоропластів, де забезпечують автономний синтез білків в цих клітинних органоїдах. Мінімальна для даного виду кількість ДНК містять статеві клітки, що мають галоїдний набір хромосом. У ядрах соматичних клітин ДНК, як правило, удвічі більше, ніж відповідає диплоїдному набору. Окрім водневих зв'язків стабілізація спіральної структури ДНК досягається також міжплощинними взаємодіями основ. Параметри моделі Уотсона - Крика відповідають конформації ДНК у фізіологічних умовах (т.з. В-форма ДНК). Нагрівання, зміна рН, пониження іонної сили, ряд інших чинників викликають денатурацію дволанцюгової молекули ДНК.
Процес реплікації (самоподвоєння) ДНК
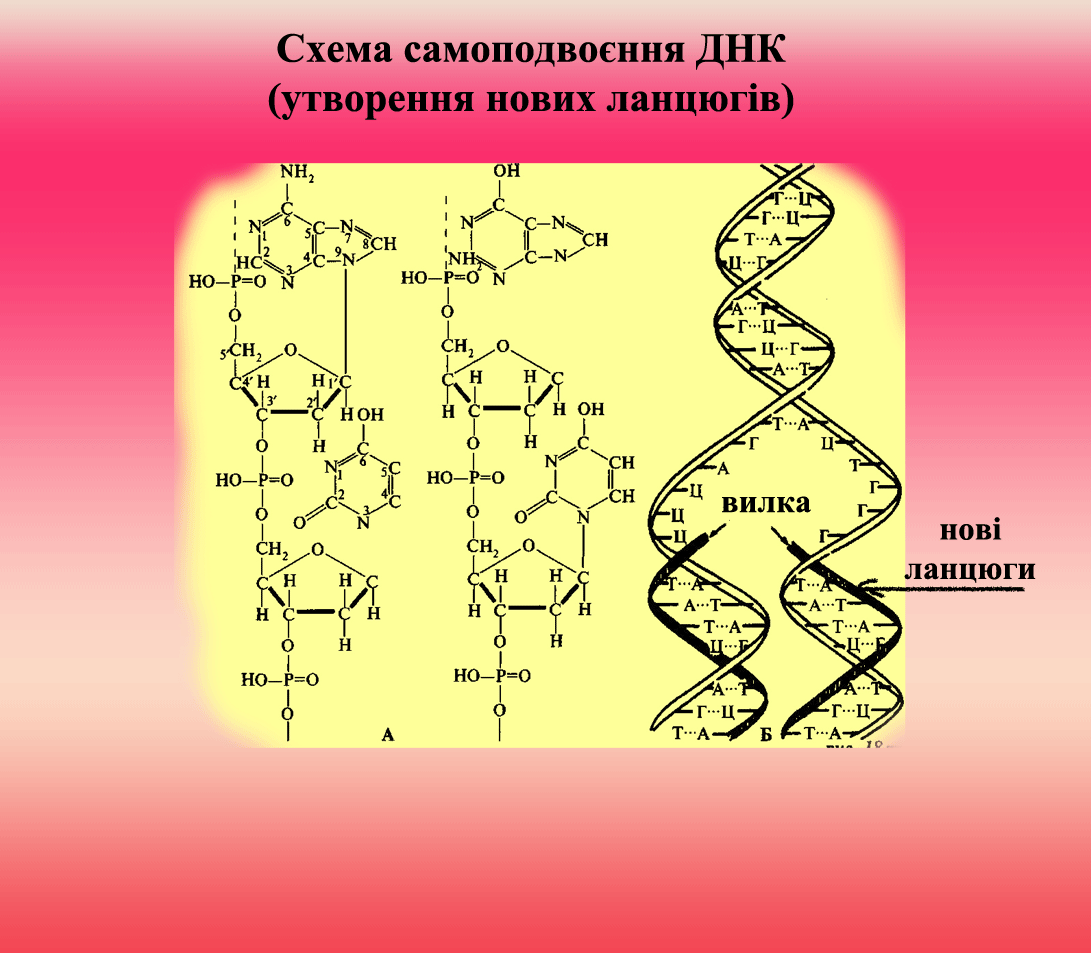
відбувається за допомогою ферментів геліказ, які розривають водневі зв’язки між ланцюгами та ДНК-полімераз, які утворюють комплементарний новий ланцюг на матриці (одному з існуючих ланцюгів). Реплікація хромосомної ДНК в клітині, що ділиться, починається з локального розплітання подвійної спіралі і утворення реплікативної вилки, в чому беруть участь специфічні ендонуклеази що розплітають білки. Синхронність реплікації обох антипаралельних ланцюгів забезпечується завдяки тому, що синтез йде короткими фрагментами (100-10 000 нуклеотідов), які приєднуються потім до ланцюгів. А. Корнберг в 1967 здійснив ферментативний синтез біол. активної ДНК in vitro.
Процес транскрипції – зчитування інформації з ДНК на РНК (рибонуклеїнову кислоту) відбувається також за допомогою геліказ і РНК-полімераз. При цьому формуються інформаційна РНК (і - РНК) – яка несе інформацію про відповідну послідовність амінокислот в протеїнах, складається з 300 – 30000 нуклеотидів, транспортна РНК (т-РНК) – яка несе інформацію за допомогою триплету (трьох нуклеотидів) про відповідну амінокислоту, складається з 70 – 90 нуклеотидів, рибосомна РНК – яка забезпечує певне просторове розташування іРНК і тРНК на рибосомі, становить близько 85 % загальної кількості РНК клітини і 60 % маси рибосоми.
ІІ. Функції нуклеїнових кислот, які несуть генетичну інформацію.
Роль ДНК полягає в збереженні інформації про будову організму (в першу чергу білків, їх співвідношення та розташування в організмі). Ця інформація специфічна для кожного біологічного виду.
Роль і.РНК – це перенос генетичної інформації від зони її збереження (тобто від ДНК) до міста перетворення інформації в біомолекули (білки). В процесі перетворення інформації в біомолекули приймають участь також т. РНК – молекула, яка підносить до місць синтезу білків амінокислоти і р.РНК – яка складає приблизно 60 % рибосоми – органели, на якій відбувається біосинтез білків.
Таким чином існують три види нуклеїнових кислот:
1. і. РНК - інформаційна;
2. т. РНК - транспортна;
3. р. РНК - рибосомна.
ІІІ. Будова нуклеїнових кислот, які не несуть генетичну інформацію.
ІV. Функції нуклеїнових кислот, які не несуть генетичну інформацію.
АТФ - нуклеїнові кислоти, які не несуть генетичної інформації, але виконують функцію накопичення енергії під час утворення хімічних зв’язків своїх молекул. АТФ – аденозінтрифосфорна кислота, АДФ – аденозіндифосфорна кислота, АМФ – аденозінмонофосфорна кислота (рис. 3).
АТФ, АДФ, АМФ – нуклеотиди, моно, ди, або три фосфорні ефіри аденіну. Мають в складі аденін, рибозу, один або два, або три залишки фосфорної кислоти.
АТФ ресинтезуються у еукаріотів переважно у мітохондріях, відновлюючись з АМФ та АДФ. У прокаріотів відновлення АТФ відбувається переважно у клітинній стінці.
НАД (НІКОТИНАМІДАДЕНІНДИНУКЛЕОТИД) динуклеотид, що складається з аденина, аміда нікотинової кислоти, двох залишків рибози і двох залишків фосфорної кислоти; кофермент дегідрогенази, виявлений у всіх живих клітинах, що функціонує на початкових етапах биологічного окислення жирів, білків і вуглеводів. Відкритий в 1904 в дріжджовому соку А. Гарденом і У. Йонгом; будова установлена в 1936 О. Варбургом і X. Эйлером-Хельпіном. С дегідрогеназами у реакціях, що каталізують, НАД і його фосфоріловане похідне НАДФ - проміжні акцептори і переносники електронів і водню. У клітинах НАД присутній в значно більших кількостях, чим НАДФ. У більшості тканин біосинтез НАД здійснюється багатоферментною системою як з нікотином, так і з нікотинової кіслоти; у печінці і нирках містяться ферментні системи, здатні синтезувати НАД з триптофану. НАДФ відрізняється від НАД наявністю третього залишку фосфорної кислоти. У клітинах НАДФ присутній в основному у відновленій формі (НАДФ.Н). Окислений НАДФ+ - акцептор водню при окисленні глюкозо-6-фосфату в пентозному циклі, в світлових реакціях фотосинтезу і т. і. Відновлений НАДФ Н використовується головним чином у біосинтезах жирних кислот, вуглеводів (у темнових реакціях фотосинтезу), відновному амінуванні альфа-кетоглутарової кислоти. При дії специфичної НАДФ-цитохромредуктази відбувається пряме окислення НАДФ у дыхальному ланцюзі. Біосинтез НАДФ здійснюється при фосфори-луванні НАД ферментом НАД-кіназою.






