Лекция 9. Клітинні мембрани. План. Клітинні мембрани та органели: хімічний склад, будова і функції. Транспорт речовин через мембрани. Поверхневий апарат клітини, його функції та особливості будови. Клітинну мембрану (оболонку рослин) вперше зміг побачити у звичайний мікроскоп, який сам розробив, Антуан Левенгук (див. Лекцію 1). Його мікроскоп був на перший погляд примітивним, але дозволяв збільшувати ті об’єкти у 300 разів.
Поверхневий апарат: 1. Капсула (зовнішня оболонка), яка складається на 98 % з водин і на 2 % з білків. Виконує захисну функцію. 2.
Клітинна стінка бактерій, специфічна по хімічному складу оболонка, що
оточує протопласт і тісно зв'язана структурно-функціональними взаєминами
з цитоплазматичною мембраною. Товщ. 10-50 нм. Складає 10-50% сухої маси
клітин. У більшості бактерій в склад входить опорний полімер
пептідоглікан (муреїн). У грампозитивних бактерій пептідоглікан може
складати до 95% клітинної стінки. У невеликій кількості до
грампозитивних бактерій входять тейхоєві і тейхуронові к-ти,
полісахариди і білки. Полісахариди і тейхоєві к-ти бактерій володіють
антигенними властивостями. До грамнегативних бактерій окрім тонкого
(товщ. 3-8 нм, 5-10% від сухої маси) пептідогліканового шару, звичайно у
вигляді одношарової мережі, входять зовні тришарові ліпопротєїдні
мембрани (~8 нм). Її компоненти (гліколіпіди) обумовлюють антигенні
властивості клітин, а також їх акцепторну специфічність по відношенню до
фагів і бактеріоцинів. 3. Цитоплазматична мембрана (подібна до інших клітин) Внутрішні структури: 4. Рибосоми, які складаються з двох судодиниць ( константа седиментації S70 у поєднаному стані). 5. Нуклеоїд (циклічна ДНК). 6. РНК. 7.
Плазміди - фрагментарні не основні ДНК - слугують засобом мутації й
адаптацій до змін умов навколишнього середовища. Є причиною
пристосування хвороботворних бактерій до антибіотиків. 8. Пілі, фімбрії - засоби прикріплення до субстрату. 9. Джгутики (не завжди) - засоби пересування. 10. Цитоплазма. Основні
форми бактерій представлені переважно одноклітинними організмами. Але
деякі види прокаріотів після поділу клітин утворюють різні по формі
групи, які складаються з деяких клітин. Основні форми мікроорганізмів,
які відносяться до класу Bacteria, мають циліндричну, або сферичну, або
війчасту форму. КЛІТИННА СТІНКА бактерій, специфічна по хімічному
складу оболонка, що оточує протопласт і тісно зв'язана
структурно-функціональними взаєминами з цитоплазматичною мембраною.
Товщ. 10-50 нм. Складає 10-50% сухої маси клітин. У більшості бактерій в
склад входить опорний полімер пептідоглікан (муреїн). У грампозитивних
бактерій пептідоглікан може складати до 95% клітинної стінки. У
невеликій кількості до грампозитивних бактерій входять тейхоєві і
тейхуронові к-ти, полісахариди і білки. Полісахариди і тейхоєві к-ти
бактерій володіють антигенними властивостями. До грамнегативних бактерій
окрім тонкого (товщ. 3-8 нм, 5-10% від сухої маси) пептідогліканового
шару, звичайно у вигляді одношарової мережі, входять зовні тришарові
ліпопротєїдні мембрани (~8 нм). Її компоненти (гліколіпіди) обумовлюють
антигенні властивості клітин, а також їх акцепторну специфічність по
відношенню до фагів і бактеріоцинів.
Рис. 1. Будова клітинної стінки
Цитоплазма бактерійної клітини обмежена від клітинної стінки тонкою напівпроникною структурою товщиною 5-10 нм, званою цитоплазматичною мембраною (ЦПМ). ЦПМ складається з подвійного шару фосфоліпідів, пронизаних білковими молекулами. З ЦПМ пов'язані багато ферментів і білки, що беруть участь в транслокації живильних речовин, а також ферменти і переносники електронів стадій біологічного окислення (дегідрогеназа, цитохромна система, АТФ-аза). На ЦМП локалізуються ферменти, що каталізують синтез пептидоглікану, білків клітинної стінки, власних структур. Мембрана є також місцем перетворення енергії при фотосинтезі, окислювальному фосфорилуванні.
Морфологічні структури клітин еукаріот.
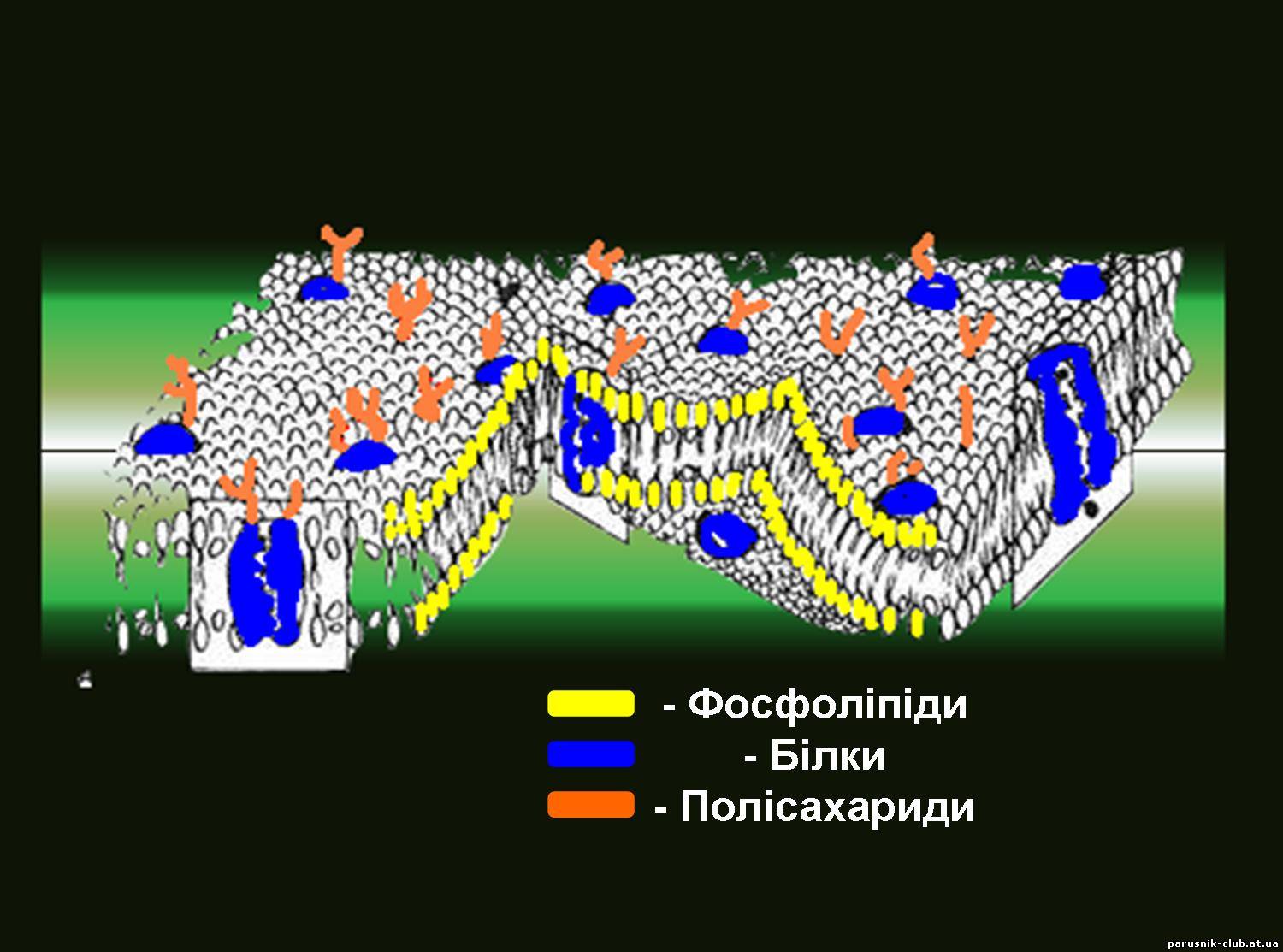
Глікокалікс – зовнішній шар еукаріотичних клітин – складається з глюкопротеїдів, виконує функцію зв’язку клітин з зовнішнім середовищем та приймає подразнення. Плазматична мембрана - існує у прокаріотів і еукаріотів. Складається з подвійного шару (бішару) фосфоліпідів (на 50 %), білків (50 %). Виконує функції: обмежувальну, опорну, пересувну.
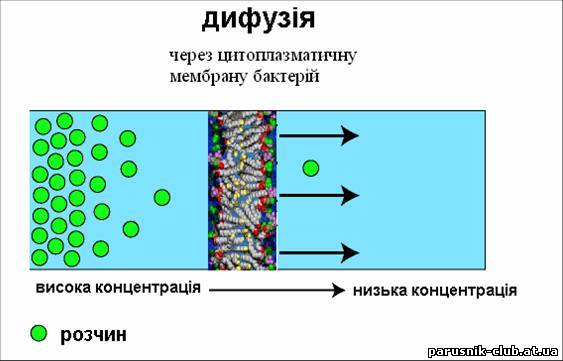
Рис. 2. Цитоплазматична мембрана. ТРАНСПОРТ РЕЧОВИН (від лат. trans-porto - переношу, переміщаю, перевожу), в живих організмах включає доставку необхідних з'єднань до певних органів і тканин (з допомогою кровоносної системи у тварин і провідної системи у рослин), всмоктування їх клітинами і пересування усередині клітин, а також виведення продуктів обміну речовин. Ці процеси можна розділити по їх механізму на транспорт - дифузію в розчинах Дифузія – процес проникнення речовин через мембрану (її пори та певні ділянки) внаслідок хаотичного теплового руху молекул без затрат енергії.
Рис. 3. Проста дифузія.
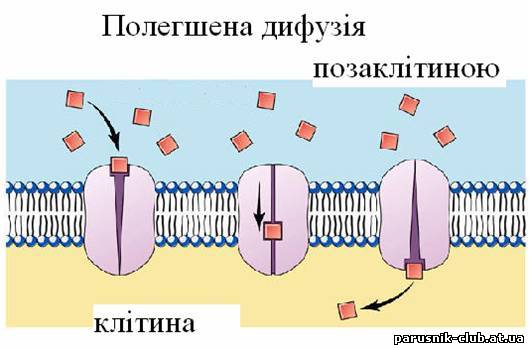
Пасивний і активний Т. в. через мембрани. У першому випадку Т. в. відбувається мимоволі, при цьому молекули і іони переносяться в область з більш низьким електрохимічним потенціалом. Пасивний Т. в. через мембрани кількісно характеризується величиною проникності, яка може різко розрізнятися для різних речовин, але кінець кінцем визначається законами дифузії і електродифузії. Проста дифузія легко відбувається через ліпідний шар мембран тільки тих речовин, що добре розчиняються в ліпідах, до яких відносяться багато ліків. Іони (Na+, К+ і Ca2+) переносяться через мембрани нервових, м'язових і ін. клітин завдяки наявності у них іонних каналів - білків (полегшена дифузія), які відкриваються і закриваються залежно від величини різниці електричних потенціалів на мембрані або дії химічних медіаторів.
Рис. 4. Полегшена дифузія.
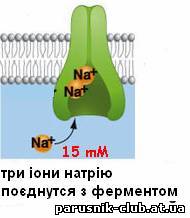
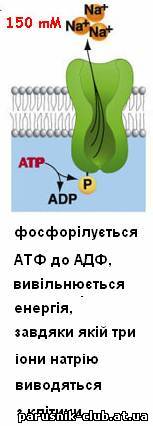
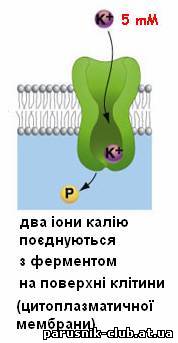
Виключення або різка зміна властивостей переносників і каналів лежить в основі дії більшості токсичних речовин. Дія ряду ліків, препаратів засновано на зміні властивостей каналів і переносників, яка дозволяє регулювати Т.. у клітинах і цілому організмі. Перенесення молекул (іонів) у зворотному напрямі (активний транспорт) можливий тільки при одночасній витраті енергії, джерелом якої може служити гідроліз АТФ в ланцюгах перенесення електронів, і здійснюється специфічними системами - іонними насосами.
Рис. 5. Активний транспорт речовин через мембрани (етапи).
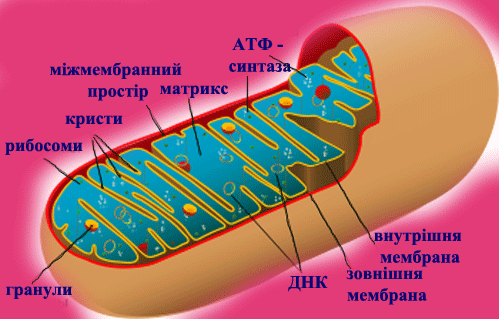
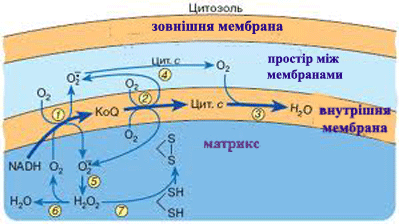
Слідством такого активного Т. р., наз. первинним, є нерівноважний розподіл іонів Н+, Са2+ Na+, K+ усередині клітини і між клітиною і позаклітинним середовищем, що у свою чергу, забезпечує роботу систем зв'язаного, або вторинного активного Т. р. через мембрани. Прикладом зв'язаного т. р. служить перенесення цукрів і амінокислот в клітини кишкового епітелію. МАТРИКС (лат. matrix, від mater - основа, букв, мати) в цитології, основна гомогенна або тонкозерниста речовина клітини, що заповнює внутріклітинні проміжки між структурами. Склад його у різних структур значно розрізняється по білках, метаболітам, іонам. Найважливіша роль М. ядра (каріоплазми), як внутрішнього напіврідкого середовища, полягає в об'єднанні всіх структур в єдину систему і забезпеченні взаємодії між ними в ядерних процесах. МІТОХОНДРІЯ (від грецького mi'tos - нитка chondrion - зернятко), органоїд еукаріотной клітини, що забезпечує організм енергією. М. описані Р. Альтманом в 1894 під назв. біобластів, а в 1897 К. Бенда назвав їх мітохондріями. Довжина варіює, досягаючи 10 мкм і трохи більш, діам. у середньому 0,2-1 мкм. Число М. в клітині коливається в широких межах - від 1 до 100 тис. М. складається з матриксу, оточеного внутрішньою мембраною, міжмембранного простору і зовнішніх мембран. У матриксі містяться кільцеві молекули мітохондріальної ДНК, специфічної іРНК, тРНК і рибосоми (прокаріотного типу), відмінні від цитоплазматичних; часто зустрічаються гранули солей кальцію і магнію; тут відбувається автономний біосинтез білків, що входять у внутрішню мембрану М., а також окислення і синтез жирних к-т. Внутрішня мембрана складається в основному з білків (70% ) фосфоліпідів (тільки 20% ) і інше; вона утворює вгинання (гребені), або трубчасті вирости - кристи, володіє строго специфічною проникністю і системами активного транспорту. Зовнішня мембрана -гладенька, товщ. 6-7 нм, складається з білків (15%) і фосфоліпідів (85%), володіє неспецифічною проникністю для більшості речовин з мол. м. менше 10 000. Осн. функція М.- утворення енергії (приблизно 95% в тваринній клітині і трохи менше в рослинній і клітині грибів). Первинна форма накопичення енергії - електрохімічний потенціал, що виникає на внутрішній мембрані. Б. ч. його відразу ж витрачається на синтез АТФ, частина безпосередньо використовується на активний транспорт через мембрану (напр., іонів кальцію) або на вироблення тепла. Джерело енергії у М.- процеси окислення біологічного (тканинного, або клітинного, дихання), що починаються з окислення піровиноградної к-ти, тієї, що утворюється в цитоплазмі при гліколізі. Перший етап - розщеплювання пірувату і реакції циклу трікарбонових к-т - відбувається в матриксі. Другий етап - перенесення електронів від водню по ланцюгу дихальних ферментів на кисень і синтез АТФ, тобто окислювальне фосфоріліруваня - у внутр. мембрані. Электрохи-міч. потенціал виникає в результаті роботи ланцюга перенесення електронів Системи активного перенесення у внутр. мембрані забезпечують транспорт всередину М. іонів К+ Ca2+, Mg2+, концентрація яких в матриксі М. значно вище, ніж в цитоплазмі. Ультраструктура М. сильно залежить від фізіол. стани клітини і організму, різко міняється під впливом дихальних отрут. Виділені М., як правило, пошкоджені і дуже чутливі до складу середовища. Збільшення числа М. в клітині відбувається звичайно в результаті їх ділення шляхом перешнуровування. М. відсутні в клітинах анаеробних організмів, напр, у кишкових амеб. Згідно однієї з гіпотез, М. відбулися з аеробних бактерій в результаті симбіогенеза.
Рис. 6. Мітохондрія і робота мітохондріального комплекса мембран.
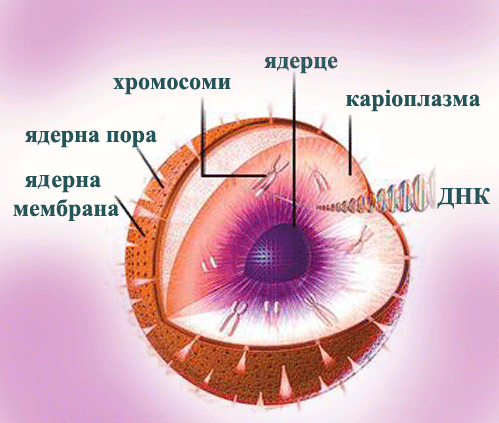
ЯДЕРНА
ОБОЛОНКА, кариолемма (karyolemma), структура, що відмежовує ядро клітин
еукаріот від цитоплазми. Складається з 2 паралельних ліпопротєїдних
мембран товщ. 7-8 нм кожна, між ними - перинуклеарний простір. Я. о.
пронизана ядерними порами діам. 60-100 нм, на кінцівках яких зовнішня
мембрана Я. о. переходить у внутрішню. Число пір від одиниць до 200 на 1
мкм1 поверхні ядра. Кожна пора по краю несе кільце щільної речовини
(аннулус), а в її просвіті звичайно є центр, елемент діам. 15-20 нм,
сполучений з аннулусом радіальними фібрілламі; ці структури складають
поровий комплекс, який регулює проходження макромолекул (білків
рібонуклеопротєїдів і ін.) через пори. Наруж. мембрана місцями може
тимчасово переходити в мембрани ендоплазматчної сітки, звичайно вона
несе рібосоми, внутрішня - часто вистілана зсередини шаром волокнистої
речовини (ядерна пластинка, або ламіна). Ядро: складається з поверхневого апарату та внутрішнього середовища – матрикса. Ядерце,
нуклеола (nucleolus), щільне тільце усередині ядра більшості клітин
еукаріот. Складається з рибонуклеопротеїдів (РНП) - попередників
рибосом. Звичайно в ядрі є одне Я., рідше декілька або багато (напр., в
ядрах яйцеклітин риб, що ростуть). Я. формується на відповідних локусах
хромосом (ядерцевих організаторах), де знаходяться серії генів, що
кодують рибосомну РНК (рРНК). Рідше (особливо в яйцеклітинах, а також в
макронуклеусах інфузорії) Я. утворюється на зовні-хромосомних копіях
ядерцевого організатора. Я. складається із зони всередині - ядерцевого
хроматину, зони фібрілл РНП товщ. 5-10 нм (що містять знов синтезовані
молекули прерібосомной РНК з константою седіментації 45 S) і зони гранул
діам. 10-20 нм (звичайно на периферії) - попередників великих і малих
субодиниць рибосом, що відповідно містять молекули рРНК з константами
седіментації 28 S і 18 S. Прерибосомні гранули відділяються від Я. і
мігрують в цитоплазму, де і відбувається збірка рибосом. На
світломікроскопічному рівні фіблярна зона Я. описується як аморфна
частина, а гранулярна - як нуклеолонема (сітчаста гетерогенна частина).
При мітозе Я. звичайно розпадається, а після закінчення його формується
наново.
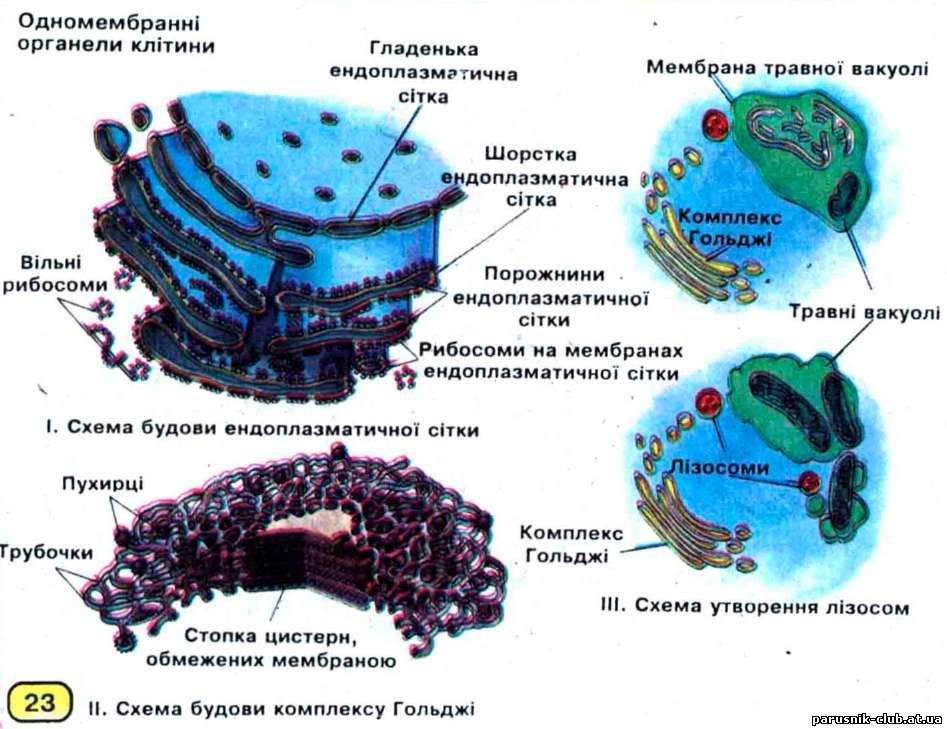
ЕНДОПЛАЗМАТИЧНА СІТКА, ендоплазматичний ретикулум (від ендо... і плазма), органоїд еукаріотной клітини. Відкритий К. Портером в 1945 році в ендоплазмі фібробластів. Ендоплазматична сітка є системою дрібних вакуолей і канальців, сполучених один з одним і обмежених одинарною мембраною. Мембрани Е. с., товщ. 5-7 нм, у ряді випадків безпосередньо переходять в зовнішню ядерну мембрану. Похідними Е.с. є мікротільця, а в рослинних клітинах - вакуолі. Розрізняють гладеньку (агранулярную) і гранулярну Е. с. Гладенька Е. с. позбавлена рибосом. Складається з канальців і дрібних вакуолей діаметром 50-100 нм. Мабуть, є похідним гра- нулярной Е. с., в деяких випадках їх мембрани безпосередньо переходять один в одного. Функції: синтез трігліцеридів і ліпідів клітин, накопичення крапельок ліпідів, обмін деяких полісахаридів (глікоген), накопичення і виведення з клітини отруйних речовин, синтез стероїдних гормонів. У м'язових волокнах утворює саркоплазматична сітка. Гранулярна Е. с. має рибосоми на мембранах. Складається з канальців і цистерн, формує розгалужену мережу. Основна функція - синтез білків на прикріплених до мембрани зовні комплексах рибосом - полірибосомах. Синтезуються в основному білки, які виводяться з клітини назовні або трансформуються в комплексі Гольджі. Синтезіровані білки поступають в порожнини гранулярної Е. с., де здійснюється АТФ-залежний транспорт білків і може відбуватися їх модифікація і концентрація. Найбільш розвинена в клітинах з білковою секрецією (підшлункової залози, слинних залоз, плазмоцитах і т. п.) і практично відсутня в ембріональних недиференційованих клітинах.
Рис. 7. Ядерна структура й принцип роботи ядра (передача генетичної інформації до ЕПС)
КОМПЛЕКС ГОЛЬДЖІ, апарат Гольджі, пластинчастий комплекс (complexus lamellosus), клітинний органоїд, що виконує ряд важливих функцій. Відкритий К. Гольджі (1898) в нервових клітинах. За допомогою електронної мікроскопії було показано, що присутній у всіх еукаріотичних клітинах. Будова його в різних клітинах сильно варіює. Структурно-функціональна одиниця К. Г.- діктіосома. У клітині міститься до 20 (рідко більш) діктіосом, розподілених в цитоплазмі дискретно або зв'язаних в загальну мережу. Область К. Г. практично позбавлена рибосом, в тваринних клітинах вона часто оточує центріолі. К. Г. розташовується в апікальной частині клітини і в його склад входять секреторні гранули, що формуються. Функції К. Г.: модифікація білків (глюкозіліруваня, сульфатування, фосфорілування, часткове розщеплювання поліпептидних ланцюгів і т. п.), упаковка ферментів в гранули (лізосоми), синтез деяких полісахаридів, формування клітинної мембрани, утворення лізосом. Білки поступають в К. Г. з гранулярної ендоплазматичної сітки в мембранних бульбашках. У К.Г. з них утворюються складні білки (ліпопротєїди, мукопротєїди, мукопо-лісахариди). Готові продукти накопичуються в бульбашках і потім безпосередньо включаються в мембрану.