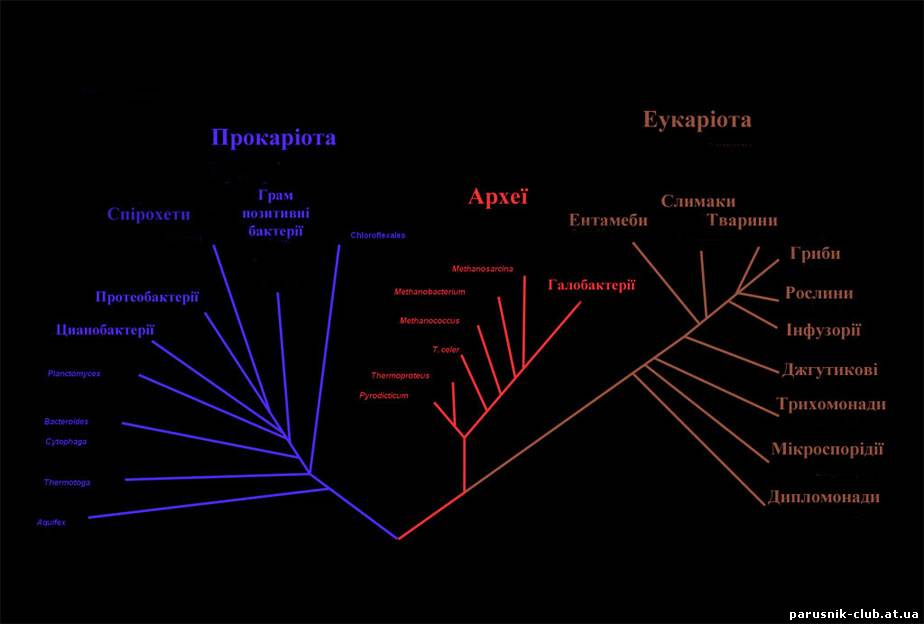
Доядерні або прокаріоти (Prokaryotes, від давньогрецького pro- перед + karyon горіх або ядро, посилаючись на ядро клітини + суфікс -otos, pl. -otes; також може писатися як "procaryotes") - організми без ядра (= karyon) клітини, (у більшості випадків також без будь-яких інших мембранних органел та одноклітинні, але є винятки). Цей набір характеристик відрізняє їх від еукаріотів (ядерних організмів), які мають клітинні ядра і можуть бути як одноклітинними, так і багатоклітинними. Різниця між структурою прокаріотів та еукаріотів найбільша серед груп організмів. Більшість прокаріотів - бактерії, і ці два терміни раніше розглядалися як синоніми. Проте, американський вчений Карл Воуз запропонував поділ прокаріотів на бактерій і архей (Bacteria та Archaea, спочатку Eubacteria і Archaebacteria) через істотні генетичні відмінності між цими групами. Система поділу на еукаріотів, бактерій та архей зараз вважається визнаною та називається Системою трьох доменів. Вона замінила собою Систему двох імперій. До прокаріот відносять: спірохети, протеобактерій, грампозитивних бактерій, цианобактерій тощо, до архей: метаносарцин, метанококусов, метанобактерій тощо, до еукаріот: найпростіших, гриби, рослин, тварин.
Рис. 1. Система трьох доменів.
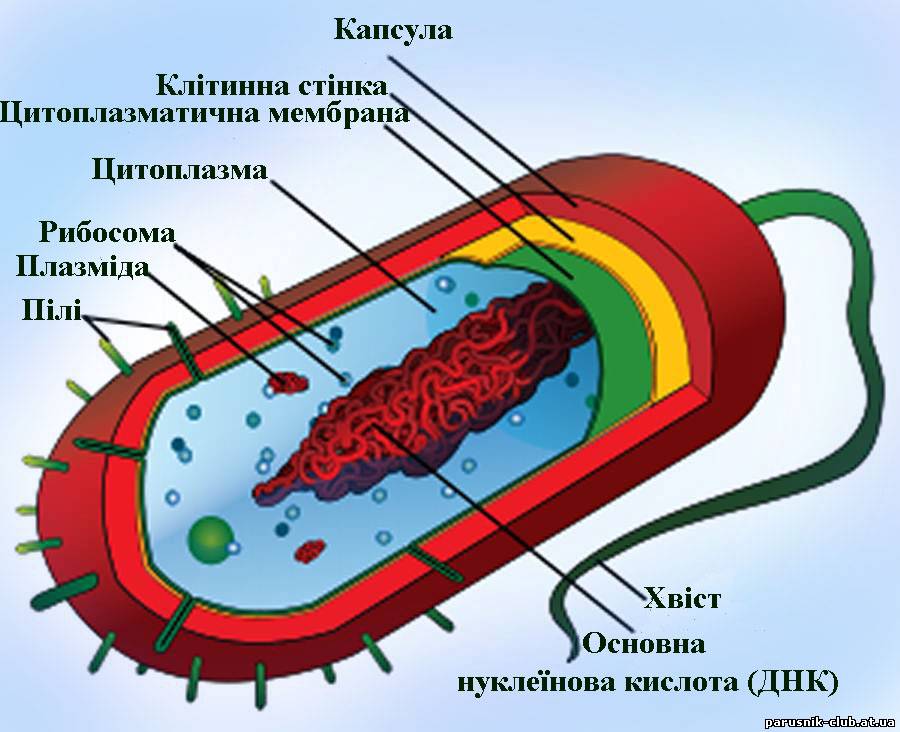
Прокаріота. Основні форми, розміри, особливості будови мікроорганізмів (бактерії, мікоплазми, рикетсії, хламідії). Будова прокаріот простіша за інши групи організмів. Вона включає (рис. 2):
Рис. 2. Будова клітини прокаріота.
Поверхневий апарат: 1. Капсула (зовнішня оболонка), яка складається на 98 % з водин і на 2 % з білків. Виконує захисну функцію. 2. Клітинна стінка бактерій, специфічна по хімічному складу оболонка, що оточує протопласт і тісно зв'язана структурно-функціональними взаєминами з цитоплазматичною мембраною. Товщ. 10-50 нм. Складає 10-50% сухої маси клітин. У більшості бактерій в склад входить опорний полімер пептідоглікан (муреїн). У грампозитивних бактерій пептідоглікан може складати до 95% клітинної стінки. У невеликій кількості до грампозитивних бактерій входять тейхоєві і тейхуронові к-ти, полісахариди і білки. Полісахариди і тейхоєві к-ти бактерій володіють антигенними властивостями. До грамнегативних бактерій окрім тонкого (товщ. 3-8 нм, 5-10% від сухої маси) пептідогліканового шару, звичайно у вигляді одношарової мережі, входять зовні тришарові ліпопротєїдні мембрани (~8 нм). Її компоненти (гліколіпіди) обумовлюють антигенні властивості клітин, а також їх акцепторну специфічність по відношенню до фагів і бактеріоцинів (рис. 10). 3. Цитоплазматична мембрана (подібна до інших клітин)
Внутрішні структури: 4. Рибосоми, які складаються з двох судодиниць ( константа седиментації S70 у поєднаному стані). 5. Нуклеоїд (циклічна ДНК). 6. РНК. 7. Плазміди - фрагментарні не основні ДНК - слугують засобом мутації й адаптацій до змін умов навколишнього середовища. Є причиною пристосування хвороботворних бактерій до антибіотиків. 8. Пілі, фімбрії - засоби прикріплення до субстрату. 9. Джгутики (не завжди) - засоби пересування. 10. Цитоплазма. Основні форми бактерій представлені переважно одноклітинними організмами. Але деякі види прокаріотів після поділу клітин утворюють різні по формі групи, які складаються з деяких клітин. Основні форми мікроорганізмів, які відносяться до класу Bacteria, мають циліндричну, або сферичну, або війчасту форму.
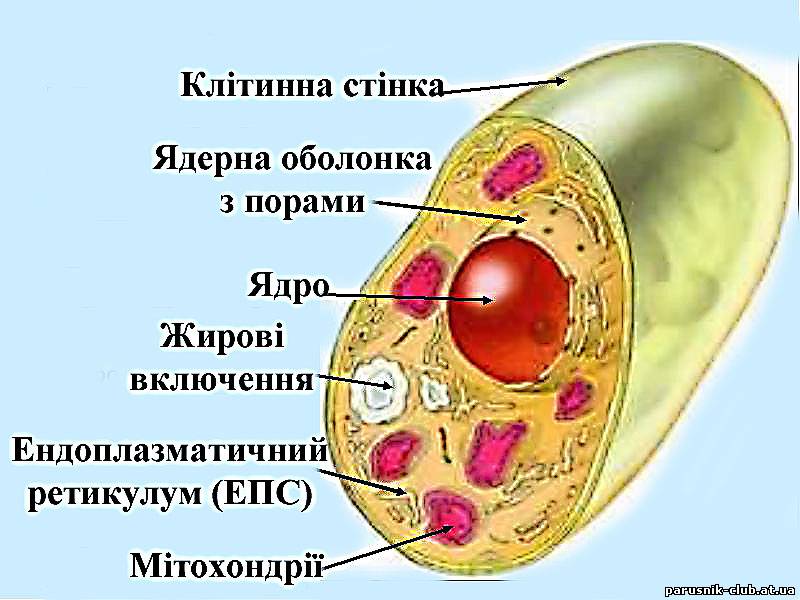
Еукаріо́ти (інколи Евкаріо́ти, від грец. еу -повністю, добре й каріон - ядро) або Я́дерні (Eukaryota Whittaker & Margulis, 1978) — домен одно- та багатоклітинних організмів, що характеризуються переважно полігеномними клітинами, морфологічно сформованим ядром та наявністю мембранних субклітинних органел (рис. 3).
Рис. 3. Будова клітини евкаріота.
Геноми еукаріотичної клітини представлені: а) ядерним геномом, зосередженим у ядрі і представленим ядерною ДНК; б) У більшості клітин мітохондріальним геномом, зосередженим у мітохондрії і представленим мітохондріальною ДНК; в) У деяких клітин пластидним (зокрема хлоропластним) геномом, що розташовується у пластиді, і представлений хлоропластною ДНК (генофором); г) геномом нуклеоморфу[1]. , який виявлений лише у кількох відділах водоростей у надзвичайно цікавій ядроподібній структурі, розташованій між оболонкою пластиди та особливою клітинною системою — хлоропластним ендоплазматичним ретикулумом. В нуклеоморфі виявлена власна, нуклеоморфна ДНК. Двогеномні клітини, в яких представлені ядерний та мітохондріальний геноми, характерні для грибів та тварин; тригеномні — з ядерним, мітохондріальним та пластидним геномами — для майже всіх рослин; чотиригеномні — з ядерним, мітохондріальним, пластидним та нуклеоморфними геномами, виявлені у хлорарахніофітових та криптофітових водоростей, а також у деяких видів з відділу Dinophyta.
В цій лекції буде розглянуто тільки деякі структури і органели клітин, які відіграють основну роль у підтримки життєдіяльності клітини. До таких структур відносяться:
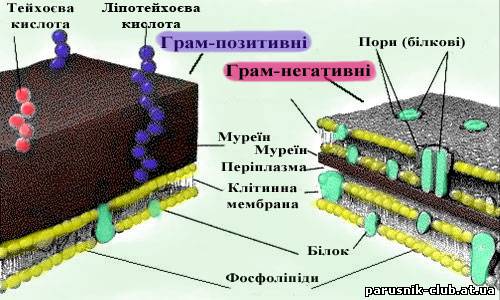
Поверхневий апарат прокаріот та еукаріот. Капсула – зовнішня желеподібна оболонка бактерій. Складається з ліпополісахаридів. Глікокалікс – зовнішній шар еукаріотичних клітин – складається з глюкопротеїдів, виконує функцію зв’язку клітин з зовнішнім середовищем та приймає подразнення. Клітинна стінка бактерій, специфічна по хімічному складу оболонка, що оточує протопласт і тісно зв'язана структурно-функціональними взаєминами з цитоплазматичною мембраною. Товщ. 10-50 нм. Складає 10-50% сухої маси клітин. У більшості бактерій в склад входить опорний полімер пептідоглікан (муреїн).
Рис. 4. Клітинна стінка бактерій.
У грампозитивних бактерій пептідоглікан може складати до 95% клітинної стінки. У невеликій кількості до грампозитивних бактерій входять тейхоєві і тейхуронові к-ти, полісахариди і білки. Полісахариди і тейхоєві к-ти бактерій володіють антигенними властивостями. До грамнегативних бактерій окрім тонкого (товщ. 3-8 нм, 5-10% від сухої маси) пептідогліканового шару, звичайно у вигляді одношарової мережі, входять зовні тришарові ліпопротєїдні мембрани (~8 нм). Її компоненти (гліколіпіди) обумовлюють антигенні властивості клітин, а також їх акцепторну специфічність по відношенню до фагів і бактеріоцинів. У грибів (рис. 5) - клітинна стінка відносно тонка, представлена полісахаридом хітином. У рослинній клітині (рис. 6) клітинна стінка також існує, але представлена полісахаридом целюлозою, деякими іншими сполуками, які надають їй пружності.
Рис. 5. Будова клітини гриба.
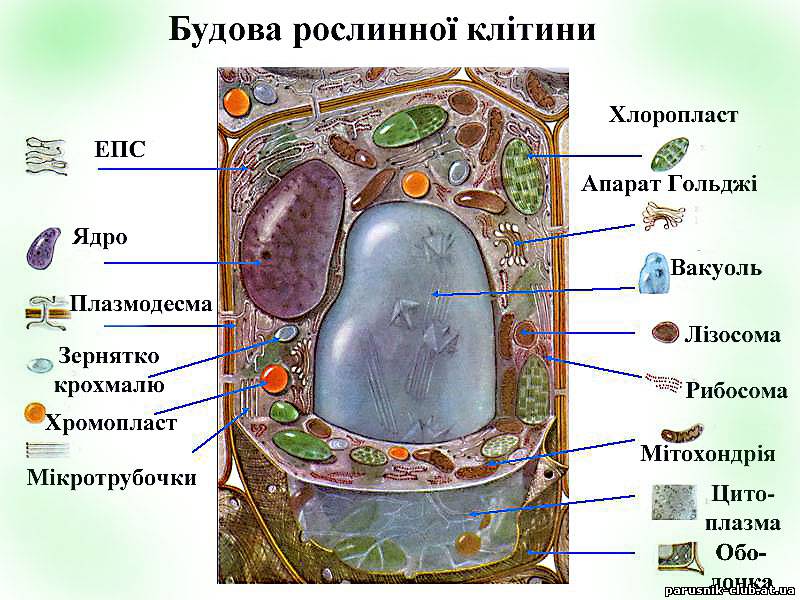
Рис. 6. Будова клітини рослин.
Плазматична мембрана - існує у прокаріотів і еукаріотів. Складається з подвійного шару (бішару) фосфоліпідів (на 50 %), білків (50 %). Виконує функції: обмежувальну, опорну, пересувну. Ця тема буде детальніше розглянута у наступній лекції.
Транспорт речовин (від лат. trans-porto - переношу, переміщаю, перевожу), в живих організмах включає доставку необхідних з'єднань до певних органів і тканин (з допомогою кровоносної системи у тварин і провідної системи у рослин), всмоктування їх клітинами і пересування усередині клітин, а також виведення продуктів обміну речовин. Ці процеси можна розділити по їх механізму на транспорт - дифузію в розчинах Дифузія – процес проникнення речовин через мембрану (її пори та певні ділянки) внаслідок хаотичного теплового руху молекул без затрат енергії. Пасивний і активний Т. в. через мембрани. У першому випадку Т. в. відбувається мимоволі, при цьому молекули і іони переносяться в область з більш низьким електрохимічним потенціалом. Пасивний Т. в. через мембрани кількісно характеризується величиною проникності, яка може різко розрізнятися для різних речовин, але кінець кінцем визначається законами дифузії і електродифузії. Проста дифузія легко відбувається через ліпідний шар мембран тільки тих речовин, що добре розчиняються в ліпідах, до яких відносяться багато ліків. Іони (Na+, К+ і Ca2+) переносяться через мембрани нервових, м'язових і ін. клітин завдяки наявності у них іонних каналів - білків (полегшена дифузія), які відкриваються і закриваються залежно від величини різниці елек-тричних потенціалів на мембрані або дії химічних медіаторів. Виключення або різка зміна властивостей переносників і каналів лежить в основі дії більшості токсичних речовин. Дія ряду ліків, препаратів засновано на зміні властивостей каналів і переносників, яка дозволяє регулювати Т.. у клітинах і цілому організмі. Перенесення молекул (іонів) у зворотному напрямі (активний транспорт) можливий тільки при одночасній витраті енергії, джерелом якої може служити гідроліз АТФ в ланцюгах перенесення електронів, і здійснюється специфічними системами - іонними насосами. Слідством такого активного Т. р., наз. первинним, є нерівноважний розподіл іонів Н+, Са2+ Na+, K+ усередині клітини і між клітиною і позаклітинним середовищем, що у свою чергу, забезпечує роботу систем зв'язаного, або вторинного активного Т. р. через мембрани. Прикладом зв'язаного т. р. служить перенесення цукрів і амінокислот в клітини кишкового епітелію.
Ядерна оболонка, кариолемма (karyolemma) (далі Я.о.), структура, що відмежовує ядро клітин еукаріот від цитоплазми. Складається з 2 паралельних ліпопротєїдних мембран товщ. 7-8 нм кожна, між ними - перинуклеарний простір. Я. о. пронизана ядерними порами діам. 60-100 нм, на кінцівках яких зовнішня мембрана Я. о. переходить у внутрішню. Число пір від одиниць до 200 на 1 мкм1 поверхні ядра. Кожна пора по краю несе кільце щільної речовини (аннулус), а в її просвіті звичайно є центр, елемент діам. 15-20 нм, сполучений з аннулусом радіальними фібрілламі; ці структури складають поровий комплекс, який регулює проходження макромолекул (білків рібонуклеопротєїдів і ін.) через пори. Наруж. мембрана місцями може тимчасово переходити в мембрани ендоплазматчної сітки, звичайно вона несе рібосоми, внутрішня - часто вистілана зсередини шаром волокнистої речовини (ядерна пластинка, або ламіна).
Ядро: складається з поверхневого апарату та внутрішнього середовища – матрикса. Матрикс (лат. matrix, від mater - основа, букв, мати) (далі М.) в цитології, основна гомогенна або тонкозерниста речовина клітини, що заповнює внутріклітинні проміжки між структурами. Склад його у різних структур значно розрізняється по білках, метаболітам, іонам. Найважливіша роль М. ядра (каріоплазми), як внутрішнього напіврідкого середовища, полягає в об'єднанні всіх структур в єдину систему і забезпеченні взаємодії між ними в ядерних процесах.
Ядерце, нуклеола (nucleolus) (далі Я.), щільне тільце усередині ядра більшості клітин еукаріот. Складається з рибонуклеопротеїдів (РНП) - попередників рибосом. Звичайно в ядрі є одне Я., рідше декілька або багато (напр., в ядрах яйцеклітин риб, що ростуть). Я. формується на відповідних локусах хромосом (ядерцевих організаторах), де знаходяться серії генів, що кодують рибосомну РНК (р.РНК). Рідше (особливо в яйцеклітинах, а також в макронуклеусах інфузорії) Я. утворюється на зовні-хромосомних копіях ядерцевого організатора. Я. складається із зони всередині - ядерцевого хроматину, зони фібрілл РНП товщ. 5-10 нм (що містять знов синтезовані молекули прерибосомної РНК з константою седиментації 45 S) і зони гранул діам. 10-20 нм (звичайно на периферії) - попередників великих і малих субодиниць рибосом, що відповідно містять молекули р.РНК з константами седіментації 28 S і 18 S. Прерибосомні гранули відділяються від Я. і мігрують в цитоплазму, де і відбувається збірка рибосом. На світломікро-ськопічному рівні фіблярна зона Я. описується як аморфна частина, а гранулярна - як нуклеолонема (сітчаста гетерогенна частина). Під час мітозу Я. звичайно розпадається, а після закінчення його формується наново.
Хромосоми (від хромо... і сома) (далі Х.), органоїди клітинного ядра, що є носіями генів і що визначають спадків, властивості клітин і організмів. Здібні до само відтворення, мають структурну і функціональну індивідуальність і зберігають її в ряду поколінь. Термін "Х." запропонований В. Вальдейером (1888). Основу X. складає одна безперервна дволанцюжкова молекула ДНК (у X. ок. 99% ДНК клітини), пов'язана з білками (гистонами і ін.) у нуклеопротеїд. Будовою молекули ДНК, її генетіч. кодом забезпечується запис спадкової інформації в X., білки (у X. вищих рослин і тварин їх міститься до 65% ) беруть участь в складній упаковці ДНК в X. і регуляції її здібності до синтезу РНК - транскрипції. В процесі функціонування X. зазнають структурно-морфологічних перетворень, в основі яких лежить процес спіралізації - деспіралізації структурних субодиниць X.- хромонем. На стадії метафази ділення клітин спіралізовані (щільноупаковані) хромосоми добре помітні в світловому мікроскопі. Кожна X. складається з двох подовжніх копій - хроматид, що утворилися в ході редуплікації і скріплені центромерою. У клітинах тіла двостатевих тварин і рослин кожна X. представлена двома т.з. гомологічними X., що з’являються одна від материнського, а інша від батьківського організму. Статеві клітки що утворилися в результаті мейозу, містять тільки одну з двох гомологичних X. Число X. сильно варіює: від двох до сотень X. складають хромосомні набори (див. Каріотип) різних видів. Кожен вид організмів володіє характерним і постійним набором хромосом в клітині, закріплений в еволюції даного виду, а його зміни відбуваються тільки в результаті мутацій.
Ендоплазматична сітка, ендоплазматичний ретикулум (від ендо... і плазма), органоїд еукаріотной клітини. Відкритий К. Портером в 1945 році в ендоплазмі фібробластів. Ендоплазматична сітка є системою дрібних вакуолей і канальців, сполучених один з одним і обмежених одинарною мембраною. Мембрани Е. с., товщ. 5-7 нм, у ряді випадків безпосередньо переходять в зовнішню ядерну мембрану. Похідними Е.с. є мікротельця, а в рослинних клітинах - вакуолі. Розрізняють гладеньку (агранулярную) і гранулярну Е. с. Гладенька Е. с. позбавлена рибосом. Складається з канальців і дрібних вакуолей діам. 50-100 нм. Мабуть, є похідним гра- нулярной Е. с., в деяких випадках їх мембрани безпосередньо переходять один в одного. Функції: синтез трігліцеридів і ліпідів клітин, накопичення крапельок ліпідів, обмін деяких полісахаридів (глікоген), накопичення і виведення з клітини отруйних речовин, синтез стероїдних гормонів. У м'язових волокнах утворює саркоплазматична сітка. Гранулярна Е. с. має рибосоми на мембранах. Складається з канальців і цистерн, формує розгалужену мережу. Основна функція - синтез білків на прикріплених до мембрани зовні комплексах рибосом - полірибосомах. Синтезуються в основному білки, які виводяться з клітини назовні або трансформуються в комплексі Гольджі. Синтезовані білки поступають в порожнини гранулярної Е. с., де здійснюється АТФ-залежний транспорт білків і може відбуватися їх модифікація і концентрація. Найбільш розвинена в клітинах з білковою секрецією (підшлункової залози, слинних залоз, плазмоцитах і т. п.) і практично відсутня в ембріональних недиференційованих клітинах.
Комплекс Гольджі, апарат Гольджі (далі К.г.), пластинчастий комплекс (complexus lamellosus), клітинний органоїд, що виконує ряд важливих функцій. Відкритий К. Гольджі (1898) в нервових клітинах. За допомогою електронної мікроскопії було показано, що присутній у всіх еукаріотичних клітинах. Будова його в різних клітках сильно варіює. Структурно-функціональна одиниця К. г.- діктіосома. У клітині міститься до 20 (рідко більш) діктіосом, розподілених в цитоплазмі дискретно або зв'язаних в загальну мережу. Область К. г. практично позбавлена рибосом, в тваринних клітинах вона часто оточує центріолі. К. г. розташовується в апікальній частині клітини і в його склад входять секреторні гранули, що формуються. Функції К.г.: модифікація білків (глюкозолірування, сульфатування, фосфорілування, часткове розщеплювання поліпептидних ланцюгів і т. п.), упаковка ферментів в гранули (лізосоми), синтез деяких полісахаридів, формування клітинної мембрани, утворення лізосом. Білки поступають в К. г. з гранулярної ендоплазматичної сітки в мембранних бульбашках. У К.г. з них утворюються складні білки (ліпопротєїди, мукопротєїди, мукополісахариди). Готові продукти накопичуються в бульбашках і потім безпосередньо включаються в мембрану.
Мітохондрія (отгреч. mi'tos - нитка chondrion - зернятко, крупинка) (далі М.), органоїд еукаріотной клітини, що забезпечує організм енергією. М. описані Р. Альтманом в 1894 під назв. біобластів, а в 1897 К. Бенда назвав їх мітохондріями. Довжина варіює, досягаючи 10 мкм і трохи більш, діам. у середньому 0,2-1 мкм. Число М. в клітині коливається в широких межах - від 1 до 100 тис. М. складається з матриксу, оточеного внутрішньою мембраною, міжмембранного простору і зовнішніх мембран. У матриксі містяться кільцеві молекули мітохондріальної ДНК, специфічної і.РНК, т.РНК і рибосоми (прокаріотного типу), відмінні від цитоплазматичних; часто зустрічаються гранули солей кальцію і магнію; тут відбувається автономний біосинтез білків, що входять у внутрішню мембрану М., а також окиснення і синтез жирних к-т. Внутрішня мембрана складається в основному з білків (70% ) фосфоліпідів (тільки 20% ) і інше; вона утворює вгинання (гребені), або трубчасті вирости - кристи, володіє строго специфічною проникністю і системами активного транспорту. Зовнішня мембрана - гладенька, товщ. 6-7 нм, складається з білків (15%) і фосфоліпідів (85%), володіє неспецифічною проникністю для більшості речовин з мол. м. менше 10 000. Основна функція М.- утворення енергії (приблизно 95% в тваринній клітині і трохи менше в рослинній і клітині грибів). Первинна форма накопичення енергії - електрохімічний потенціал, що виникає на внутрішній мембрані. Більша частина його відразу ж витрачається на синтез АТФ, частина безпосередньо використовується на активний транспорт через мембрану (напр., іонів кальцію) або на вироблення тепла. Джерело енергії у М.- процеси окислення біологічного (тканинного, або клітинного дихання) матеріалу, що починаються з окиснення піровиноградної к-ти, тієї, що утворюється в цитоплазмі при гліколізі: Перший етап - розщеплювання пірувату і реакції циклу трикарбонових к-т - відбувається в матриксі. Другий етап - перенесення електронів від водню по ланцюгу дихальних ферментів на кисень і синтез АТФ, тобто окислювальне фосфорілування - у внутрішній мембрані. Електрохімічний потенціал виникає в результаті роботи ланцюга перенесення електронів. Системи активного перенесення у внутрішній мембрані забезпечують транспорт всередину М. іонів К+ Ca2+, Mg2+, концентрація яких в матриксі М. значно вище, ніж в цитоплазмі. Ультраструктура М. сильно залежить від фізіологічного стану клітини і організму, різко міняється під впливом дихальних отрут. Виділені М., як правило, пошкоджені і дуже чутливі до складу середовища. Збільшення числа М. в клітині відбувається звичайно в результаті їх ділення шляхом перешнуровування. М. відсутні в клітинах анаеробних організмів, напр, у кишкових амеб. Згідно однієї з гіпотез, М. відбулися з аеробних бактерій в результаті симбіогенезу. Ленінджер А., Мітохондрія, пер. з англ.. М., 1966.
Хлоропла́сти — тип пластиди, органела, знайдена в клітинах рослин і деяких водоростей, що не відносяться до рослин. Хлоропласти поглинають сонячне світло і використовують його разом з водою та вуглекислим газом для отримання енергії для рослини (у формі АТФ) шляхом фотосинтезу. Назва цієї органели походить від грецького слова chloros — «зелений» і plast, що посилається на форму органели і може бути перекладеним як «органела» або «клітина». Хлоропласти — одні з унікальних структур в організмі рослини і, як загалом вважається, виникли в результаті ендосимбіозу від ціанобактерій. У цьому відношенні вони подібні до мітохондрій, але знайдені тільки в рослинах та найпростіших. Обидві органели оточені складною подвійною мембраною з міжмембранним простором, мають свою власну ДНК і залучені до енергетичного метаболізму, обидві мають багато виступів на своїй внутрішній мембрані. У зелених рослинах хлоропласти оточені двома мембранами. Вважається, що внутрішня мембрана хлоропласту відповідає зовнішній мембрані спадкової ціанобактерії. Геном хлоропласту значно скорочений порівняно з геномом вільно-живучих ціанобактерій, але існуючі частини вказують на спільність походження. Багато з відсутніх генів кодуються в ядерному геномі клітини-хазяїна. У деяких водоростях (наприклад гетероконтах) та інших найпростіших (наприклад Euglenozoa і Cercozoa), хлоропласти, здається, з'явилися через вторинний ендосимбіоз, в якому еукаріотична клітина охопила другу еукаріотичну клітину, що містить хлоропласти, формуючи хлоропласти з трьома або чотирма мембранами. У деяких випадках такі вторинні ендосимбіонти містять у собі іншу еукаріотичну клітину, формуючи третинний ендосимбіоз. Хлоропласти — плоскі диски зазвичай 2-10 мікрометра в діаметрі і 1 мікрометр завтовшки. Хлоропласт має дві мембранні оболонки — внутрішню і зовнішню мембрани. Між ними знаходиться міжмембранний простір. Рідина у хлоропласті, строма, відповідає цитозолю бактерій і містить дуже маленьку кільцеву ДНК і рибосоми, хоча більшість їх білків кодуються генами ядра клітини, продукція яких транспортується до хлоропласту. В межах строми знаходяться багато шарів тилакоїдів, суб-органел, де фактично протікає фотосинтез. Декілька шарів тілакоїдів разом називаються граном. Тілакоїд нагадує плоский диск, усередині якого — порожня область, що називається тілакоїдним простором або люменом. Реакція фотосинтезу протікає на тілакоїдній мембрані і, як і в мітохондріях, використовує по'єднання мембранного транспорту з біосинтезом. У телакоїдній мембрані знаходяться диски молекул хлорофілу, антенні комплекси. Вони допомогають збільшити площу, з якої захоплюється світло. Потім енергія фотонів направляється до центру комплексу. Дві молекули хлорофілу іонізуються на фотон захопленого світла, виробляючи збуджені електрони, які потім потрапляють до фотосинтетичного реакційного центру. Хімічний склад хлоропластів представлений каротиноїдами - 4,5%, білками - 35-55%, ліпідами - 20-30%, вуглеводами - 10%, РНК - 2-3%, ДНК - 0,5% та хлорофілом - 9%(С55Н72О5N4Mg) В хлоропластах сконцентровані 80% Fe, 70% Zn, близько 50% Si.