Пластичний обмін речовин (анаболізм, або асиміляція) - це сукупність фізіолого-біохімічних процесів, в ході яких з простих органічних і неорганічних речовин утворюються складніші речовини. Пластичний обмін протікає з витратою високоорганізованої енергії (наприклад, у вигляді АТФ), яка витрачається на відновлення початкових сполук вуглецю шляхом приєднання до них електронів і протонів. Для пластичного обміну потрібні первинні джерела вуглецю (початкова "цегла" для утворення органічних речовин) і первинні джерела високоорганізованої енергії та тепла. Усі організми здатні синтезувати складні органічні речовини, використовуючи відносно прості органічні речовини з асиметричним атомом вуглецю. Організми, усі клітини яких потребують готових органічних речовин, називаються гетеротрофними (чи просто гетеротрофами). Проте існують організми, у яких частина клітин здатна асимілювати (тобто засвоювати) вуглекислий газ. Такі організми називаються автотрофними (чи просто автотрофами). До них відносяться рослини й синьо-зелені водорості. До автотрофів часто відносять й деяких інших прокаріот, здатних до автотрофного (хемоавтотрофного, де використовується не енергія світла, а енергія хімічних сполук ) засвоєння речовин. Усі організми здатні отримувати високоорганізовану енергію шляхом катаболізму (тобто за рахунок окислення органічних речовин). Організми, у яких усі клітини отримують високоорганізовану енергію тільки таким шляхом, називаються органотрофними (чи просто органотрофами). Проте існують організми, у яких хоч би частина клітин здатна використати світлову енергію. Термодинаміка живих систем. Другий закон термодинаміки оворить, що ентропія (міра невпорядкованості) в ізольованій системі постійно зростає. Іншими словами можна сказати, що безлад «наводиться сам», а для впорядкування системи потрібно виконати хімічну, механічну та транспортну роботу. Її постійно змушені виконувати організми, для того щоб підтримувати життєдіяльність, це відбувається завдяки наявності метаболізму. Системи із великим запасом вільної енергії є нестабільними, в них самовільно відбуваються процеси, що зменшують її. Отже спонтанно можуть відбуватись тільки реакції, що супроводжуються виділенням енергії (ΔG < 0), вони називаються екзергонічними, але не реакції, для яких ΔG > 0 (ендергонічні). У стані термодинамічної ΔG=0 система не може виконувати роботу. Досягнення стану рівноваги для клітини означає смерть. Якщо пряма реакція екзергонічна, як наприклад окиснення глюкози до вуглекислого газу й води ΔG0 = −686 ккал /моль (2879 кДж /моль)), то зворотна обов'язково буде ендергонічною, тобто для реакції синтезу глюкози із вуглекислого газу та води ΔG0 = +686 ккал/моль. Зміна вільної енергії під час реакції залежить від її конкретного типу (тобто вмісту енергії у вихідних речовинах і продуктах), а також від того, наскільки далеко була система від термодинамічної рівноваги у початковий момент часу (тобто наскільки концентрація речовин відрізняються від рівноважних). В реальних умовах живої клітини більшість хімічних реакцій ніколи не досягають рівноважного стану через те, що продукти постійно використовуються в інших реакціях або виділяються у середовище. Таким чином жива система перебуває у динамічно стабільному стані: у ній відбуваються неперервні зміни внаслідок току речовини та енергії, але основні показники підтримуються на сталому рівні. Наприклад глюкоза з одного боку поглинається з крові і використовується багатьма тканинами, з іншого — всмоктується із кишечника та мобілізується з місць її запасання. Внаслідок поєднання цих двох процесів її концентрація у крові залишається досить стабільною. БІОСИНТЕЗ (від біо... і греч. synthesis - з'єднання), утворення органічних речовин з простіших сполук, що відбувається в живих організмах під дією біокаталізаторів - ферментів. Б.- важлива сторона обміну речовин живих організмів. Біосинтез залежний від процесів розщеплювання складніших речовин на простіші. Джерелом енергії для Б. служать багаті енергією (макроергічних фосфатів) сполуки, а початковою (для всіх організмів окрім бактерій, що здійснюють хемосинтез) - є енергія сонячного випромінювання, акумульована зеленими рослинами і ціанооактеріямі в процесі фотосинтезу (про що було наведено раніше). Кожен одноклітинний організм як і кожна клітина багатоклітинного організму, синтезує його складові речовини. Характер Б., здійснюваного в клітині, визначається спадковою інформацією, закодованою в її генетичному апараті. Б., здійснюваний мікроорганізмами широко застосовується в біотехнології як спосіб проміжного отримання вітамінів, деяких гормонів, антибіотиків, амінокислот, а також кормових білків і ін. сполук. ФОТОСИНТЕЗ (від фото... і греч. synthesis - з'єднання), утворення клітинами вищих рослин водоростей і деякими бактеріями органічних речовин за участю енергії світла. Відбувається з допомогою пігментів (хлорофілів і інших), присутніх в хлоропластах і хроматофорах. У основі Ф. лежить окисно-відновлювальний процес, в яком електрони переносяться від донора-відновника (вода, водень і ін.) до акцептора (СО2, ацетат) з утворенням відновлених з'єднань (вуглеводи) і виділенням O2, якщо окиснюється H2O (якщо фотосинтезують бактерії, що використовують інші, чим вода, донори, кисень не виділяють). Етапи фотосинтеза представлені у таблиці 1. Таблиця 1. Етапи фотосинтезу.
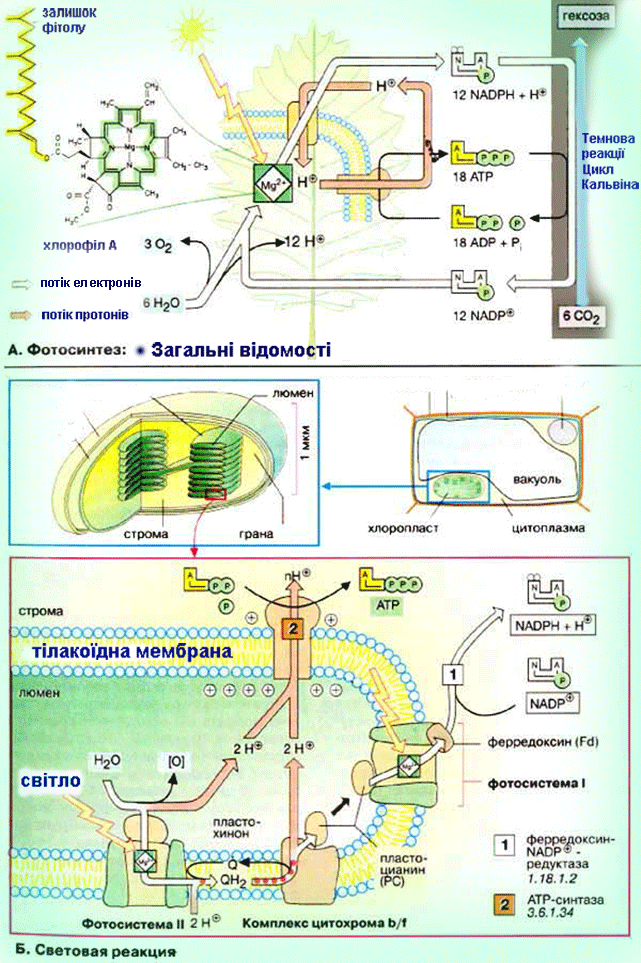
Схема двох фотохімічних систем (ФС1 і ФСII) фотосинтезу.
Перетворення енергії світла в енергію хімічних зв'язків починається в специфічних структурах - реакційних центрах (РЦ). Вони складаються з молекул хлорофілу
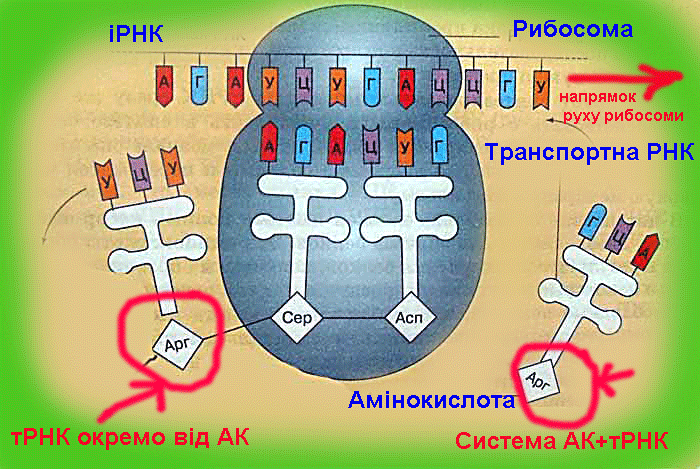
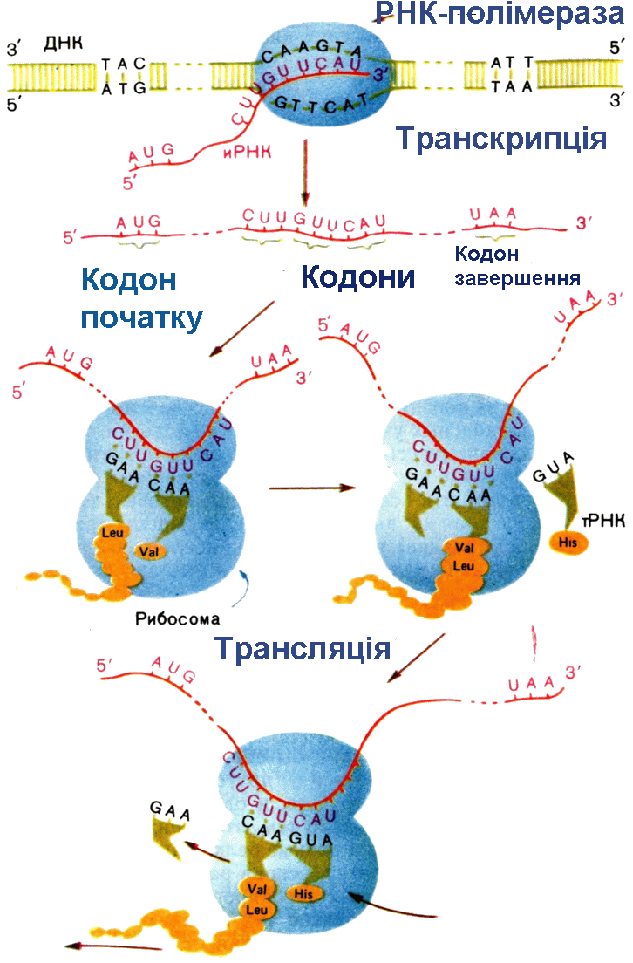
а (у бактерій - бактеріохлорофілу, у галобактерій - бактеріородопсину), виконуючих функцію фотосенсибілізаторів, пігменту феофітину, пов'язаних з ними донорів і акцепторів електронів і інших з'єднань. У Ф. вищих рослин, водоростей, ціанобактерій беруть участь дві послідовні фотореакції з різних РЦ. При поглинанні квантів пігментами фотосистеми II (ФС II) відбувається перенесення електронів від води до проміжному акцептору і через ланцюг перенесення електронів до РЦ фотосистеми I (ФС I). Збудження ФС I супроводжується перенесенням електрона на другий ступінь (через проміжний акцептор і ферредоксин до НАДФ+). У РЦ зосереджена лише невелика (~1% ) частина хлорофілу що безпосередньо бере участь в перетворенні енергії поглинених фотонів в енергію хімічних зв'язків, основна його маса і додаткові (супроводжуючі) пігменти виконують роль світлозбираючої антени. Декілька десятків або сотень таких молекул, зібраних у т.з. фотосинтетичні одиниці, поглинають кванти і передають збудження на пігментні молекули РЦ. Це значно підвищує швидкість Ф. навіть при невисоких інтенсивностях світла. У РЦ відбувається утворення первинних відновника і окислювача, які потім ініціюють ланцюг послідовних. окис.-відн. реакцій, і енергія у результаті запасається у відновленому никотінамідаденіндінуклеогидфосфаті (НАДФ.Н) і АТФ (фотосиитетіч. фосфорилування) - осн. продуктах фотохімічних світлових стадій Ф. Продукти первинних стадій Ф. вищих рослин і водоростей, в яких запасена енергія світла, використовуються надалі в циклі фіксації СО2 і перетворенні вуглецю у вуглеводи (т.з. цикл Калвіна). СО2 приєднується до рибулозодифосфату за участю ферменту рібулозодіфосфаткарбоксилази. З одержаного шестивуглецевого з'єднання утворюється трьохвуглецева (С3) фосфогліцеринова к-та (ФГК) відновлювана потім з використанням АТФ і НАДФ.Н до трьохвуглецевих цукрів (тріозофосфатов), з яких і утворюється кінцевий продукт Ф.- глюкоза. Разом з тим частина тріозофосфатів зазнає процес конденсації і перебудови перетворюючись на рибулозомонофосфат, який фосфорілуєтся за участю "свеітлового" АТФ до рибулозодіфосфату - первинного акцептора CO2, що і забезпечує безперервну роботу циклу. У деяких рослинах (кукурудза, цукровий очерет і ін.) первинне перетворення вуглецю йде не через трьохвуглецеві, а через чотирьохвуглецеві з'єднання (С4-рослини C4-метаболизм вуглецю). Акцептором СО2 в клітинах мезофіла таких рослин служить фосфоенолпіруват (ФЕП). Продукти його карбоксилювання - яблучна або аспарагинова к-ти дифундують в обкладки судинних пучків, де декарбоксилюются з звільненням СО2, який і потрапляє в цикл Кальвіна. Переваги такого "кооперативного" метаболізму обумовлені тим, що ФЕП-карбоксилаза при низькій концентрації СО2 активніша, чим рибулозодіфосфаткарбоксилаза, і, крім того, в обкладочних клітинах із зниженою концентрацією O2 слабкіше виражено фотодихання, пов'язане з окисленням рибулозофосфату, і супутні йому втрати енергії (до 50%). С4-рослини привертають увагу дослідників високої фотосинтетичною продуктивністю. Фотосинтез єдиний процес в біосфері, ведучий до збільшення вільної енергії біосфери за рахунок зовнішнього джерела - Сонця і що забезпечує існування як рослин, так і всіх гетеротрофних організмів, в т.ч. і людини. Щорічно в результаті Ф. на Землі утворюється 150 млрд. т органічної речовини і виділяється біля 200 млрд. т вільного О2. Коллобіг О2, вуглецю і ін. елементів, що залучаються у Ф., створив і підтримує сучасний склад атмосфери, необхідний для життя на Землі. Ф. перешкоджає збільшенню концентрацій СО2 в атмосфері, запобігаючи перегріву Землі (унаслідок т.з. парникового ефекту). Кисень Ф. необхідний не тільки для життєдіяльності організмів, але і для захисту живого від згубного короткохвильового УФ-випромінювання (киснево-озоновий екран атмосфери). Запасена в продуктах Ф. енергія (у вигляді видів палива) є осн. джерелом енергії для людства. Передбачається, що в енергетиці майбутнього Ф. може зайняти одне з перших місць як невичерпне джерело і таке, що не забруднює середовище (творення плантацій швидкорослих рослин з подальшим використанням рослинної маси для отримання теплової енергії або переробки у високоякісне паливо - спирт). Не менш важлива роль Ф. як основи отримання продовольства, кормів, технічної сировини. Не дивлячись на високу ефективність початкових фотофізичних і фотоімічних стадій (ок. 95%), в урожай переходить лише менше 1-2% сонячної енергії; втрати обумовлені неповним поглинанням світла, лімітацією процесу на біохімічних і фізіологічних рівнях. Забезпечення рослин водою мінеральним живленням, СО2, селекція сортів з високою ефективністю Ф., створення сприятливої структури посівів і ін. використовують в цілях реалізації резервів фотосинтетичної продуктивності. Біосинтез білка. ТРАНСКРИПЦІЯ (від лат. transcriptio, букв.- переписування), біосинтез молекул РНК, на ділянках ДНК; перший етап реалізації генетичної інформації в живих клітинах. Здійснюється ферментом ДНК залежною РНК-полімеразою, яка у більшості вивчених організмів є комплексом 4 і більш неідентичних субодиниць, що виконують різні ролі в процесі Т. Фермент «дізнається» про знак початку Т.- промотор (ділянка ДНК), приєднується до нього, розплітає подвійну спіраль ДНК і копіює, починаючи з цього місця, один з її ланцюгів, переміщаючись уздовж ДНК і послідовно приєднуючи мономірні ланки (нуклеотиди) до тієї, що утворює РНК відповідно до принципу комплементарності. У міру руху РНК-ПОЛІМЕРАЗИ ланцюг РНК, що росте, відходить від матриці і подвійна спіраль ДНК позаду ферменту відновлюється. Коли РНК-ПОЛІМЕРАЗА досягає кінця копійованої ділянки (термінатора), РНК відділяється від матриці. Число копій різних ділянок ДНК може мінятися в ході розвитку організму. Для високоефективної ініціації часто потрібне приєднання до промотора білків позитивного контролю (напр., білка-активатора катаболізму). Показано, що у прокаріот в регуляції на етапі ініціації можуть брати участь білки-репрессори, зв'язуючі близькі до крапки початку Т. ділянки - оператори. Деякі знаки кінця Т. (термінатори) пізнаються самою РНК-полімеразою, в пізнаванні ін. бере участь особливий термінуючий білок В регуляції Т. на етапі термінації беруть участь білки-антітермінатори і компоненти апарату білкового синтезу. У еукаріот існують самостійні РНК-полімерази для синтезу рибосомальних, інформац. і транспортних РНК. Одиниці Т., звані oпeронами, у прокаріот включають, як правило, декілька функціонально зв'язаних генів, у еукаріот вони завжди або майже завжди моногенни. У опухолеродних вірусів можливе перенесення информации з РНК на ДНК (зворотна Т.У за допомогою ферменту зворотної транскріптази (ревертази). ТРАНСЛЯЦІЯ (від лат. translatio - передача), синтез поліпептидних ланцюгів білків на матриці інформаційної РНК згідно генетичного коду; другий етап реалізацій генетичної інформації у живих клітинах. У. процесі трансляції інформація про специфічну будову майбутнього білка, записана у вигляді послідовності нуклеотидов в молекулах іРНК, перекладається з нуклеотидного коду у визначену послідовність амінокислот в білках, що синтезуються. Здійснюється складним макромолекулярним комплексом, що полягає окрім іРНК з рибосоми, транспортних РНК (тРНК) аміноацил-тРНК-синтетаз, білкових чинників ініціації, елонгаціі (подовження, або нарощування поліпептіду), термінаціЇ (закінчення) Т. н ін. Амінокислоти доставляються в рибосоми тРНК. На етапі ініціації Т. менша субодиниця рибосоми, ініциаторна (у бактерій формілметіонілова) тРНК і чинники ініціації "дізнаються" про кодон-ініціатор у 5'-кінца іРНК. Після цього приєднується велика субодиниця рибосоми
і в ній починається власне синтез білка, який відбувається в 3 етапи: 1. Приєднання т. РНК, 2. Утворення пептидного зв'язку 3. Просування рибосоми на три нуклеотида - транслокація.
після чого весь цикл повторюється. При пізнаванні кодонов-терминаторів білкові чинники термінації каталізують звільнення поліпептидного ланцюга від рибосоми. При синтезі білка іРНК входить до складу полірибосоми (на ній одночасно ведуть синтез до 100 рибосом). У прокаріот полірибосоми утворюються в ході транскрипції, на іРНК, ще пов'язаної з ДНК. У еукаріот синтез білка обмежений цитоплазмою.