Лекция 4.
Водный и солевой обмен у гидробионтов.
Прежде чем рассмотреть вопрос о водно-солевом обмене у гидробионтов
необходимо сказать о том, какие биофизические законы законы
задействованы в регуляции процессов такого обмена. Осмотическое давление
(обозначается π) — избыточное гидростатическое давление на раствор,
отделённый от чистого растворителя полупроницаемой мембраной, при
котором прекращается диффузия растворителя через мембрану. Это давление
стремится уравнять концентрации обоих растворов вследствие встречной
диффузии молекул растворённого вещества и растворителя.
Мера
градиента осмотического давления, то есть различия водного потенциала
двух растворов, разделённых полупроницаемой мембраной, называется
тоничностью. Раствор, имеющий более высокое осмотическое давление по
сравнению с другим раствором, называется гипертоническим, имеющий более
низкое — гипотоническим.
Осмотическое давление может быть весьма
значительным. В дереве, например, под действием осмотического давления
растительный сок (вода с растворёнными в ней минеральными веществами)
поднимается по ксилеме от корней до самой верхушки. Одни только
капиллярные явления не способны создать достаточную подъёмную силу —
например, секвойям требуется доставлять раствор на высоту до 100 метров.
При этом в дереве движение концентрированного раствора, каким является
растительный сок, ничем не ограничено.
Если же подобный раствор
находится в замкнутом пространстве, например, в клетке крови, то
осмотическое давление может привести к разрыву клеточной мембраны.
Именно по этой причине лекарства, предназначенные для введения в кровь,
растворяют в изотоническом растворе, содержащем столько хлорида натрия
(поваренной соли), сколько нужно, чтобы уравновесить создаваемое
клеточной жидкостью осмотическое давление. Если бы вводимые
лекарственные препараты были изготовлены на воде или очень сильно
разбавленном (гипотоническом по отношению к цитоплазме) растворе,
осмотическое давление, заставляя воду проникать в клетки крови,
приводило бы к их разрыву. Если же ввести в кровь слишком
концентрированный раствор хлорида натрия (3-5-10 %, гипертонические
растворы), то вода из клеток будет выходить наружу, и они сожмутся. В
случае растительных клеток происходит отрыв протопласта от клеточной
оболочки, что называется плазмолизом. Обратный же процесс, происходящий
при помещении сжавшихся клеток в более разбавленный раствор, —
соответственно, деплазмолизом.
Величина осмотического давления,
создаваемая раствором, зависит от количества, а не от химической природы
растворенных в нём веществ (или ионов, если молекулы вещества
диссоциируют), следовательно, осмотическое давление является
коллигативным свойством раствора. Чем больше концентрация вещества в
растворе, тем больше создаваемое им осмотическое давление. Это правило,
носящее название закона осмотического давления, выражается простой
формулой, очень похожей на некий закон идеального газа:
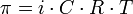
где i — изотонический коэффициент раствора;
C — молярная концентрация раствора, выраженная через комбинацию основных единиц СИ, то есть, в моль/м3.
R — универсальная газовая постоянная;
T — термодинамическая температура раствора.
Это
показывает также схожесть свойств частиц растворённого вещества в
вязкой среде растворителя с частицами идеального газа в воздухе.
Правомерность этой точки зрения подтверждают опыты Ж. Б. Перрена (1906):
распределение частичек эмульсии смолы гуммигута в толще воды, в общем,
подчинялось закону Больцмана.
Осмотическое давление, которое зависит
от содержания в растворе белков, называется онкотическим (0,03 — 0,04
атм.). При длительном голодании, болезни почек концентрация белков в
крови уменьшается, осмотическое давление в крови снижается, и возникают
осмотические отёки: вода переходит из сосудов в ткани, где πОСМ больше.
При гнойных процессах πОСМ в очаге воспаления возрастает в 2-3 раза, так
как увеличивается число частиц из-за разрушения белков. В организме
осмотическое давление должно быть постоянным (≈ 7,7 атм.). Поэтому
пациентам вводят изотонические растворы (растворы, осмотическое давление
которых равно πПЛАЗМЫ ≈ 7,7 атм. (0,9 % NaCl — физиологический раствор,
5 % раствор глюкозы). Гипертонические растворы, у которых π больше, чем
πПЛАЗМЫ, применяются в медицине для очистки ран от гноя (10 % NaCl),
для удаления аллергических отёков (10 % CaCl2, 20 % глюкоза), в качестве
слабительных лекарств (Na2SO4∙10H2O, MgSO4∙7H2O).
Закон
осмотического давления можно использовать для расчёта молекулярной массы
данного вещества (при известных дополнительных данных).
Население
морских, пересоленных, солоноватых и пресных вод имеет свои резко
выраженные особенности водно-солевого обмена. Связано это с разными
способностями противостоять гиперосмотической или гипоосмотической
среде. Чаще всего причина кроется в наследственности. Генетически
подобными являются морские и пресноводные формы, за счет которых позже
возникло население пересоленных и солоноватых вод. Среди последних нет
эндемичных семейств и рядов, в то время как среди населения морских и
пресных вод есть даже эндемичные классы и типы.
Население
пересоленных вод состоит из эвригалинных (см. на сайте страницу «Словарь
биологических терминов» в разделе «Библиотека») морских видов, живущих
при солености до 75-80‰, и форм пресноводного происхождения. Обычно
встречаются при > 80‰ (рачки Artemia salina, личинки комара
Chironomus salinarius и др..). В солоноватых водах главным образом
встречаются морские эвригалинных и пресноводные виды, а также небольшое
число специфических солоноватоводных форм. Общих форм для морей и
пресных вод практически нет, если не считать тех, которые попеременно
ведут морской или пресноводный образ жизни (проходные формы рыб, таких
как осетр европейский и т.д.). Для морских и пресноводных форм крайняя
граница распространения лежит в пределах 7-8‰. Важно отметить, что
указанная граница прослеживается в самых разных морях: Азовском и
Черном, Каспийском, Белом, Балтийском, Северном и др. А также для
различных фаунистических групп. Это может означать, что данная граница
экологической валентности по отношению к такому параметру, как соленость
воды, имеет универсальный характер. Связана такая особенность с
влиянием критической солености на многие биологические процессы.
Соленость 7-8% оказывается предельной для морских и пресноводных форм, и
вследствие этого населения вод с соленостью 7-8% о оказывается
обедненным. Флора макрофитов, по сравнению с фауной, по мере опреснения
вод обедняется не так сильно. Так, число видов в Белом море снижено на
50%, а макрофитов - на 24%, в Черном - на 75 и 50%, в Балтийском - на 95
и 80%. Большая устойчивость макрофитов к снижению солености связано с
наличием в них прочных клеточных оболочек, обеспечивающих механическое
сопротивление осмотическому поступлению воды в организм.
Интересно
отметить, что некоторые пресноводные формы выявляют наиболее сильное
количественное развитие при солености 3-5% о. В тех случаях, когда
морские формы переходят к существованию в опресненной воде, наблюдается
их измельчение. Как пример измельчения пресноводных форм в осолеводненых
водах можно привести балтийских моллюсков Theodoxus fluviatilus и
Bithynia tentaculata. Соленоводные формы мельчают с продвижением в
пресную воду, но с переходом в соленую их размеры не меняются (например,
моллюск Hydrobia ulvae).
В крайних условиях солености часто
снижается не только конечный размер гидробионтов, но также и темп их
роста. С ухудшением условий существования при переходе в пресную воду
связано, видимо, и снижение плодовитости гидробионтов. Наряду с
уменьшением плодовитости с переходом морских форм в пресную воду часто
наблюдается увеличение размеров яиц.
Солевой состав организмов
обнаруживает черты сходства с тем, который характерен для той среды, где
возникли и формировались отдельные виды. Поэтому в участках биосферы с
различным химическим составом содержание отдельных элементов в живых
организмах заметно варьирует. Созданное А.П. Виноградовым учение о
биогеохимических провинциях касается и гидросферы, хотя вода значительно
подвижнее, чем почва, и солевые градиенты в ней выражены слабее. Даже в
пределах одного Черного моря выделен ряд районов, планктон которых
характеризуется различным химическим составом. Имея определенный солевой
состав, организмы должны поддерживать его постоянство, причем
гидробионтам это значительно труднее осуществлять, чем наземным животным
и растениям. Покровы водных организмов проницаемы для ионов, поэтому
сохранение постоянства их соотношения по суммарной концентрации в теле,
окруженном водой с различным солевым составом, требует непрерывной и
интенсивной работы, Вода, имеющая «некомфортную» соленость нейтрализует
действие физико-химических сил выравнивания, непрерывно нарушает
химический гомеостаз гидробионтов. Пассивному солевому обмену, который
нарушает гомеостаз, организмы противопоставляют активный,
стабилизирующий соотношение отдельных ионов и их суммарную концентрацию.
Совершенством механизмов регуляции солевого состава, а также
выносливостью тканей к его изменениям определяется выживания
гидробионтов в водах с различным солевым составом.
Не только в
гомойосмотических (см. на сайте страницу «Словарь биологических
терминов» в разделе «Библиотека»), но и у подавляющего большинства
пойкилоосмотических гидробионтов существуют различные механизмы, которые
вопреки физико-химическим силам выравнивания поддерживают определенный
уровень общего содержания солей в жидкостях тела, и, что не менее важно,
обеспечивают стабильность концентрации в организме отдельных ионов и их
соотношение. Особенно характерен для многих гидробионтов так называемый
«натриевый насос». «Натриевый насос» - это вид активного транспорта
веществ через мембрану клеток. Немного остановимся на вопросе. что же
такое активный транспорт веществ с точки зрения кинетики. Активный
транспорт — перенос вещества через клеточную или внутриклеточную
мембрану (трансмембранный А.т.) или через слой клеток (трансцеллюлярный
А.т.), протекающий против градиента концентрации из области низкой
концентрации в область высокой, т. е. с затратой свободной энергии
организма. В большинстве случаев, но не всегда, источником энергии
служит энергия макроэргических связей АТФ.
Различные транспортные
АТФазы, локализованные в клеточных мембранах и участвующие в механизмах
переноса веществ, являются основным элементом молекулярных устройств —
насосов, обеспечивающих избирательное поглощение и откачивание
определенных веществ (например, электролитов) клеткой. Активный
специфический транспорт неэлектролитов (молекулярный транспорт)
реализуется с помощью нескольких типов молекулярных машин — насосов и
переносчиков. Транспорт неэлектролитов (моносахаридов, аминокислот и
других мономеров) может сопрягаться с симпортом — транспортом другого
вещества, движение которого по градиенту концентрации является
источником энергии для первого процесса. Симпорт может обеспечиваться
ионными градиентами (например, натрия) без непосредственного участия
АТФ. Активный перенос Na + обычно соединен с эквивалентным, но
противоположно направленным передвижением какого-нибудь «противоиона»,
конечно К + или NH4 +. За передвижение Na + ответственная Na +
к-АТФ-аза, что стимулируется ионами Na и К, и этот фермент является
векториальний, т.е. имеет пространственно направленное действие.
Натриевый насос в одних случаях освобождает организм от избытка ионов Na
+, поступающих пассивно, в других - работает в обратном направлении,
когда натрия в окружающей воде мало и организм теряет его в результате
диффузии. Например, у проходных рыб при адаптации в соленой воде, в
хлоридных клетках жабр (клетки Кейс-Вильмера) активность Na + к-АТФ-азы
возрастает многократно, транспортная система поляризована таким
образом, что Na + перекачивается наружу. При снижении концентрации Na + в
окружающей среде (переход рыб в пресную воду) также повышается
активность фермента, но транспортная система поляризуется в обратном
направлении, т.е. Na + перекачивается внутрь. Активный солевой обмен,
связан со способностью некоторых клеток захватывать ионы из воды или
выделять их из тела вопреки физико-химическим силам, свойственный как
растениям, так и животным. Большей устойчивостью к концентрации
различных ионов отличаются ткани рыб. У пресноводных гидробионтов
стабильная концентрация ионов в теле связана с их сорбцией клетками,
лежащими на поверхности тела, или в специальных экскреторных органах. У
костистых рыб захват или отдача ионов натрия происходит в результате
деятельности так называемых кейсвильмеровских клеток в жабрах. У морских
рыб эти клетки служат для экскреции избытка солей наружу, у
пресноводных - для захвата ионов из внешней среды. Изменение функции
кейсвильмеровских клеток наблюдается и у проходных рыб, идущих на нерест
из моря в пресную воду. Способность к плавающему захвату или отдачи
ионов теми же клетками, в зависимости от биологических потребностей,
известна для многих ракообразных и личинок насекомых (например,
Chironomus).
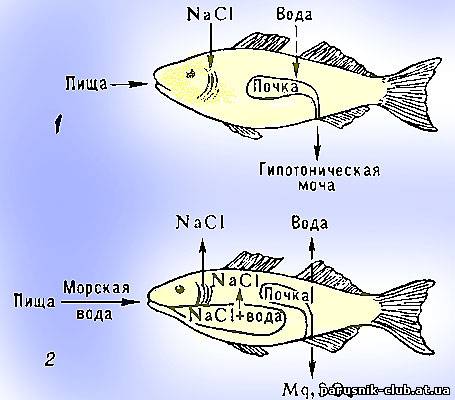
У рыб основную функцию регуляции осмотического давления в целом в организме выполняют почечные канальцы, которые реагируют своей активностью на уже случившиеся изменения гомеостаза.(см.рис).
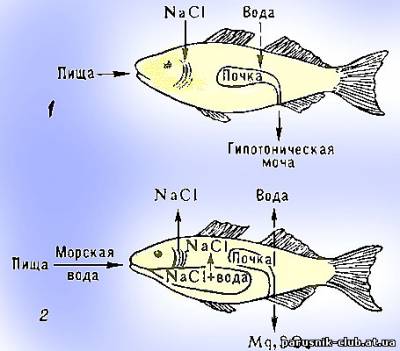
Осмотическая регуляция у рыб. А - пресноводных. Б - морских.
Однако у проходных рыб цитологические и гистохимические
преобразования в кейсвильмеровских клетках, отражающих изменения их
функции, происходят до изменения солености среды, и поэтому дальнейшее
пребывание рыбы в соленой воде становится невозможным. Повышение
осмотического давления крови, возникающее как неизбежное следствие
изменения функции клеток Кейс-Вильмера, побуждает рыбу идти в пресную
воду. Таким образом, перестройка механизма осморегуляции в этом случае
опережает будущее изменение осмотического давления среды.
Активный
захват различных ионов клетками поверхности тела может играть
существенную роль в минеральном питании многих животных. Например,
высшие раки поглощают из воды растворенный в ней кальций, цинк и другие
ионы. Через поверхность тела рыбы поглощают соединения серы, как,
например, это наблюдается в ряду карповых и осетровых, фосфор и другие
элементы минерального питания.
Постоянство солевого состава
организмов в условиях меняющейся солености воды может быть обеспечена за
счет образования осмотически непроницаемых покровов. Такой способ
защиты от осмотического обводнения и обезвоживания присущ всем водным
млекопитающим и птицам, покровы которых влаго- и соленепроницаемы.
Водным организмам, дышащим кислородом, растворенным в воде, развитию
влаго- и соленепроницаемости покровов препятствует необходимость
осуществления через них газообмен. Чем плотнее наружные покровы, тем
медленнее диффундируют через них кислород и углекислый газ, т.е.
ухудшаются условия дыхания. В связи с этим у многих животных плотность
покровов на теле в зависимости от участка тела становится весьма
различной. На участках, используемых для газообмена (жабры и т.п.),
покровы остаются тонкими и нежными, на другой поверхности тела они
утолщаются, в какой-то мере или полностью становясь влагонепроницаемыми.
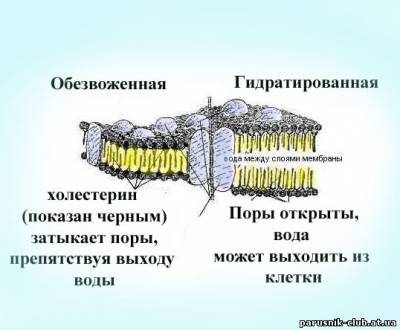
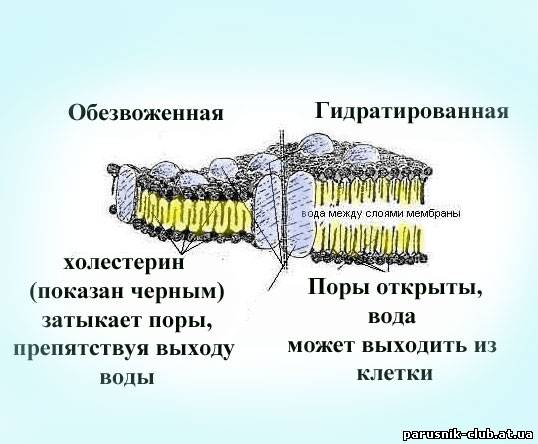
Рис. Модели сегментов насыщенной водой и обезвоженной двухслойной клеточной мембраны и канала воды, циркулирующей вокруг клетки между слоями мембраны. Канал выполняет функцию 'водного пути', где происходят все химические реакции, обеспечивающие связь клетки с внешним для нее миром.
У
ракообразных, особенно высших, кроме довольно совершенной осмотической регуляции (см. рис.)
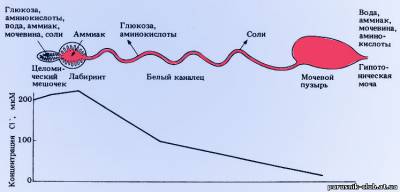
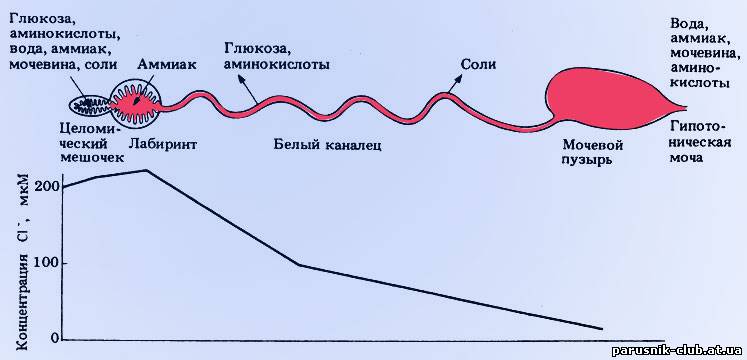
Рис. Строение и функции антенальной (зеленой) железы у раков.
существует и возможность осмотической изоляции, созданию которой способствует
развитие панциря. Сильно ослабляется отдача воды в гипертонической среде
у личинок насекомых с хорошо развитой эпи- и екзокутикулою, например у
разных стрекоз, поденок, клопов, жуков. Существенное средство
обеспечения осмотической изоляции - слизь. Например, яйца многих червей,
не способны к осморегуляции, хорошо выживают в гипо- и гипертонической
среде только потому, что заключены в слизистую кладку. Осмотическое
предназначение образования слизи на поверхности тела наблюдается у рыб и
многих других организмов. У двустворчатых моллюсков осмоизоляция в
неблагоприятной солевой среде обеспечивается смыканием раковины. Так,
мидии, устрицы и другие морские моллюски, попадая в опресненную воду,
могут долго выживать в ней с закрытыми створками, а их гемолимфа
несколько дней остается неразбавленной. Подобно моллюскам, могут
временно за счет осмоизоляции выносить резкие изменения солености
морские желуди и другие животные с раковинами, которые закрываются.
Способность многих крабов и других высших раков существовать в условиях
резких изменений солености среды в значительной мере обусловлена
водонепроницаемостью их панциря, который покрывает большую часть
поверхности тела животных. Частичная осмоизоляция облегчает работу
осморегуляторных механизмов.
У растений, живущих в гипотоническому
среде, осмотическое обводнение предупреждается прочностью клеточных
оболочек. Их механическое давление уравновешивает осмотическое и
препятствует поступлению избыточных количеств воды. Одни и те же
растения, живущие в условиях неодинаковой солености, имеют разную
тоничность, но степень их гипертонии относительно среды остается
подобной. Так, в Северном и Балтийском морях, соленость воды которых
соответственно равна 30-35 и 15-20% в, концентрация солей в
исследованных растений различалась примерно на 14% о. Какой бы ни была
соленость среды, растения создают в своих тканях чуть большее
осмотическое давление, что обеспечивает им тургор, который достигнув
определенной степени, предупреждает дальнейшее поступление воды в
клетки. Существование осмотических градиентов между тканями гидробионтов
и окружающей водой создает опасность или обезвоживания тела организмов,
или его избыточного обводнения. Чем резче осмотические градиенты, тем
эта опасность больше. Способность избегать гидратации в пресной воде и
дегидратации в морской являются основными фаторами возможной
евригалинности гидробионтов.
Среди морских организмов только
некоторые - кишечнополостные и иглокожие - имеют изотоничность к
окружающей воде. Слегка гипертонические приапулиды, кольчатые черви,
моллюски, плеченогие, большинство ракообразных; гипотоничны мизиды,
креветки и большинство крабов, а также все позвоночные. Все пресноводные
животные резко гипертоничны.
Приспособления гидробионтов к защите от
осмотического обезвоживания или обводнения прежде сводятся к
предотвращению среды с резко отличными условиями солености, как это
наблюдается у стеногалинных форм (см. на сайте страницу «Словарь
биологических терминов» в разделе «Библиотека»). У эвригалинных
гидробионтов регулирования водного обмена может достигаться частичным
или полным устранением осмотического давления за счет образования
непроницаемых покровов (осмоизоляция), сведением к минимуму разницы в
тоничности тканей и окружающей воды или противопоставлением
осмотическому давлению механического. Если эти пути не реализованы или
недостаточно эффективны, эвригалинных организмы могут существовать,
противопоставляя осмотическому давлению воды другое, направленное в
противоположную сторону и такое, которое компенсирует по своей величине
первое. Благодаря этому приток или отток воды под действием
физико-химических сил снимается физиологической работой, направленной на
сохранение водного баланса. Если защита от осмотического обезвоживания
невозможна, многие организмы, теряя влагу, впадают в анабиоз.
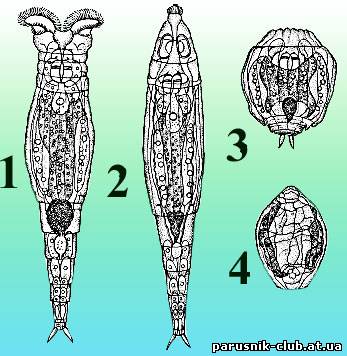
Рис. Коловратка Philidina roseola на разных стадиях высыхания (по П. Ю. Шмидту, 1948):
1– активная; 2 – начинающая сокращаться; 3 – полностью сократившаяся перед высыханием; 4 – в состоянии анабиоза.
Перекачки
организмом воды в направлении, противоположном осмотическому току,
вызывает дополнительные энергетические затраты и невыгодно. Однако и
другие защитные средства не безупречны. Уплотнение покровов, которое
ослабляет осмотическое давление, снижает диффузию через них кислорода и
СО2, то есть ухудшает условия дыхания. Сведение к нулю осмотических
градиентов за счет отождествления концентрации солей в своем теле с той,
которая имеет место в окружающей воде, невыгодно, поскольку соленость
воды варьирует, и, следовательно, гомеостаз организмов нарушается.
Поэтому выработка тех или иных средств регулирования водного обмена у
различных гидробионтов пошли в разных направлениях, обеспечивающих
наибольшую эффективность приспособлений к конкретным условиям пребывания
и биологических особенностей организмов.
В отличие от морских вод,
характеризующихся постоянством солевого состава, пресные воды различных
ландшафтных зон существенно отличаются по содержанию основных ионов.
Природные воды подразделяются по солевому составу на три класса:
1. гидрокарбонатные (С),
2. сульфатные (S),
3. хлоридные (Си).
Каждый
класс, в зависимости от преобладающих макрокомпонентов, разделяется на
три группы: кальциевая, магниевая и натриевая, а каждая группа, в свою
очередь, по количественному соотношению ионов, - на четыре типа (I, II,
III, IV).
Воды типа I образуются в процессе химического вымывания
вулканических пород или при обменных процессах Са2 + и Mg2 + на Na +.
Эти воды зачастую мало минерализованные.
Воды типа II - смешанные. Их
состав может быть связан как с осадочными породами, так и с продуктами
выветривания вулканических пород. К этому типу относятся воды
большинства рек, озер и подземных вод небольшой и умеренной
минерализации.
Тип III включает сильно минерализованные воды и воды,
характеризующиеся катионным обменом Na + на Са2 + или Mg2 +. Такие
свойства присущи водам океанов, морей, лиманов, реликтовых водоемов.
К
водам типа IV, не содержащие НСО3-, относятся кислые воды. Это воды
болот, шахтные, вулканические или воды, сильно загрязнены промышленными
стоками.
Характеристики вод сказываются следующим образом: класс -
химическим символом соответствующего аниона (С, S, C1), группа -
символом катиона (Са, Na, Mg). Принадлежность к типу сказывается римской
цифрой в нижнем индексе, в группу - символом в верхнем индексе.
Например, ССап гидрокарбонатной класс, группа кальция, тип II; ClMgIII -
хлоридный класс, группа магния, тип III и т.д.
Поверхностные воды
суши по солевому составу характеризуются такими показателями, как общая
минерализация, соотношение ионов и содержание хлоридов и сульфатов. На
большей части Европейского континента воды рек имеют небольшую или
среднюю минерализацию и относятся преимущественно к гидрокарбонатному
классу, кальциевой группы. Для степных и полупустынных зон характерна
повышенная минерализация вод сульфатного класса. В Европе такие реки
занимают лишь 3-4% площади всех речных бассейнов. Еще меньше речных
бассейнов, воды которых относятся к хлоридному классу, натриевой группы.
Как правило, эти воды характеризуются высокой минерализацией. На
территории Украины речные воды относятся преимущественно к карбонатному
классу. Основными ионами солевого состава воды Днепра и его водохранилищ
является гидрокарбонаты и Са2 +, то есть вода характеризуется
гидрокарбонатным классом, группой кальция, вторым типом.
Минерализация
и содержание отдельных ионов в воде водоемов зависят от сезона года. В
пик весеннего половодья минерализация воды в реках снижается вследствие
поступления снеговых вод. После окончания наводнения содержание солей
повышается. Рост солености воды становится ощутимым особенно в летнюю
межень и зимой, когда в питании реки увеличивается доля грунтовых вод.
Формирование
солевого состава зарегулированных вод зависит не только от состава
воды, поступающей из водосборной площади, но и от характера
внутриводоемных процессов. При многолетнем регулировании стока
минерализация воды может повышаться только до определенного уровня, то
есть до установления солевого равновесия. Сезонные колебания
минерализации и ионного состава воды крупных водохранилищ обусловлены,
главным образом, притоком речных вод, а при каскадном их расположении -
поступлением воды из вышележащих водохранилищ и незарегулированных
участков реки. В небольших водохранилищах в формировании ионного состава
воды важную роль играет также смыв растворенных солей из прибрежных
склонов, поступления грунтовых вод и атмосферных осадков, испарения,
забор воды для хозяйственно-бытовых нужд.
В озерах солевой состав
воды и ее минерализация зависят от их зонального расположения.
Соленоводные озера чаще всего расположены в степных районах юга Украины и
Крыма, т.е. в зонах с сухим, жарким климатом и высокой интенсивностью
испарения воды. В связи с этим в непроточных или слабопроточных озерах в
результате испарения происходит концентрирование основных ионов и
частичное их выпадение в осадок - в солоноватоводных озерах уже при
солености 2-10‰ и выше. В осадок переходят преимущественно
слаборастворимые карбонатные соли кальция в форме СаСО3, CaMg (CO3) 2 и
CaSО4 • 2H2O (гипс). Иногда образование таких озер связано с выходом на
поверхность грунтовых вод с повышенной минерализацией.
В воде соленых
озер концентрация солей близка к океанической или превышает ее. При
этом если нижняя граница их солености лимитируется верхним пределом
солености солоноватых (30% о) вод, то верхняя не ограничена ничем.
Соленые озера часто рассматриваются как лечебные с
высококонцентрированными рассолом, основными компонентами которых
являются Na +, K +, Са2 +, Mg2 +, СИ-, SO42-, НСО3-и CO32-. Для таких
озер характерны высококонцентрированные растворы нескольких солей, и
поэтому их класс может быть определен не по одному, а по двум анионам,
например хлоридно-сульфатный или сульфатно-хлоридный. В таком случае в
названии класса на первом месте ставится анион, который преобладает.
Выделяют следующие классы вод соленых озер: карбонатные (содовые),
сульфатные и хлоридные. В карбонатных преобладает Na2CO3, в сульфатных -
Na2SO4 и MgSO4, а в хлоридных - NaCl, MgCl2 и СаС12. В Украине есть
соленые озера, состав вод которых определяет их лечебное использование.
Это озера Крыма (Сасык, Сакское), Куяльницкий лиман в Одесской области
(находится в критическом состоянии из-за пересыхания), группа Славянских
озер (Репное, Вейсове, Слепное) в бассейне Северского Донца и некоторые
другие.