

Лекция 3.
Дыхание гидробионтов.
План.
1. Адаптация гидробионтов к газообмену.
2. Интенсивность газообмена.
3. Стойкость гидробионтов к дефициту кислорода.
4. Заморные явления.
Под интенсивностью дыхания понимается количество кислорода, потребляемого организмом в единицу времени на единицу массы. Количество потребленного кислорода оценивается в объемном или весовой единицах, вес выражается применительно ко всему веществу тела или его мягких частей (например, исключения веса раковины в моллюсков). Конечно, расчеты ведутся на сырую (живую) массу, реже на сухое вещество или его калорийный эквивалент. Отсутствие единой оценки количества вещества затрудняет анализ и сравнение имеющихся данных. Например, исключение из массы тяжелых скелетных образований при оценке дыхания моллюсков, усоногих раков и других завышает их обмен по сравнению с теми животными, у которых есть те же, но менее выраженные структуры, включают в общую массу (раковины рачков и панцири ракообразных, иглокожих и др..). Интенсивность дыхания организмов с различным содержанием воды сложно сравнивать. Поэтому необходимо проводить унификацию способов оценки массы организмов, количества вещества, на которую рассчитывается потребление кислорода. Большой интерес могут иметь данные об интенсивности дыхания, выраженные отношением энергетической стоимости кислорода, потребленного организмом, и энергоемкости последних. Такое отношение - хорошая характеристика метаболической активности живого вещества различных организмов и более точная мера интенсивности их газообмена.
Величина потребления кислорода характеризует уровень окислительных процессов, происходящих в организме, и, соответственно, его энергетические затраты в процессе жизнедеятельности. Сумма всех энергозатрат определяет величину обмена. Различают обмен основной, стандартный, активный и общий (средний). Под основным обменом, понимается уровень энергозатрат при полного покоя голодных организмов в условиях абиотической среды, близких к оптимальным. Стандартный обмен, близкий к основному, характеризует энергозатраты организмов с выключенной двигательной активностью в некоторых определенных условиях среды, в частности температурных. За величину активного обмена принимаются энергозатраты, связанные с обеспечением двигательной активности животных. Совокупность основного и активного обмена обозначают как общий или средний обмен. Потому что у беспозвоночных трудно не только вызвать состояние полного покоя, но и охарактеризовать это состояние, в опытах с гидробионтами обычно определяют стандартный или общий обмен. Животные содержатся в подопытных сосудах при отсутствии пищи тот или иной срок, после чего определяется суммарная величина потребляемого ими кислорода и вычисляется его среднее поглощения в единицу времени на единицу массы, т.е. средняя интенсивность дыхания.
Адаптация гидробионтов к газообмену.
Ускорение газообмена в гидробионтов, согласно законам диффузии может достигаться увеличением площади дыхательных поверхностей, истончением и разрыхлением покровов, через которые должны диффундировать газы
, созданием высокого градиента концентрации кислорода на внутренней и внешней сторонах покровов, в частности, путем приведения последних в контакт с аэрированная водой или газообразным кислородом.
Увеличение площади и газопроницаемости дыхательных поверхностей. Газообмен гидробионтов осуществляется либо через всю поверхность тела, или через его отдельные участки, превращены в специальные органы дыхания - жабры, трахеи, легкие и другие образования. Гидробионты, лишены специальных органов дыхания, обычно имеют тело с большой удельной поверхностью. Один из простейших способов ее увеличения заключается в уменьшении размера организмов. Небольшой размер характерен для простейших, коловраток, копепод, клещей и других организмов, не имеющих специальных органов дыхания. В мелких личинок комаров Chironоmus относительная величина жабр меньше, чем в крупных, удельная поверхность тела которых ниже, и на дыхательные органы ложится большая нагрузка. Икринки у рыб, живут и размножаются в озерах с пониженным содержанием кислорода, часто бывают мельче, чем у тех, которые живут и размножаются в озерах с более благоприятными условиями дыхания.
Увеличение поверхности тела часто достигается его уплощением, вытягиванием, образованием различных выростов, лопастей и т.п. С этим во многом связан, например, гидроморфоз растений, когда подводные листья расчленено значительно сильнее надводного, что находится в более благоприятных для дыхания условиях. Степень развития дыхательных поверхностей часто обнаруживает тесную зависимость от респираторных условий. У крабов, часто находятся на воздухе, где кислорода больше, наблюдается редукция числа и величины жабр. Иногда животные сами активно увеличивают дыхательную поверхность за счет изменения формы тела. Например, олигохеты в неблагоприятных условиях дыхания, сильно вытягиваясь в длину, утончаются, благодаря чему поверхность их тела увеличивается. Гидры и актинии при недостатке кислорода сильно вытягивают свое тело и щупальца, иглокожие - амбулакральные ножки.
Скорость диффузии газов определяется не только величиной дыхательной поверхности, но и толщиной покровов, через которые происходит газообмен. В силу этого истончение покровов дыхательных поверхностей представляет собой одну из основных адаптации к газообмена. Очень тонкие покровы на жабрах, в легких и других специальных образованиях, которые функционируют как органы дыхания. Если специальных органов дыхания нет, то утончаются покровы всего тела. Некоторые гидробионты способны увеличивать проницаемость наружных покровов, когда возникает необходимость усиления газообмена. Так, в соответствии с потребностями газообмена, изменяется степень проницаемости оболочек в икринок рыб за счет выделения гиалуронидазы.
Адаптация к использованию растворенного кислорода. Контакт дыхательных поверхностей с хорошо аэрированных водой, прежде всего, достигается в гидробионтов выбором местонахождения, где вода имеет достаточно высокое содержание кислорода, а также ее периодическим обновлением вблизи своего тела. В случае резкого ухудшения респираторного среды во организмов перемещаются даже в не свойственные им биотопы. Например, сувойка Vorticella nebulifera при дефиците кислорода образует заднее кольцо ресничек, отделяется от стебелька и ведет планктонный образ жизни до тех пор, пока респираторные условия у дна не изменятся на лучшие. Черви Nereis, личинки комара Chironomus и ряд других животных выползают на поверхность почвы, когда респираторные условия в его толще резко ухудшаются. Выбор мест с более благоприятными условиями дыхания наблюдается и в пелагических животных, когда они оставляют участки водоема с недостаточной для них концентрацией кислорода. Особенно ярко это явление наблюдается при замора, когда рыбы, клопы, жуки и другие животные нередко перемещаются на значительные расстояния в поисках достаточно аэрируемой воды.
Универсальная адаптация гидробионтов к аэрированию дыхательных поверхностей происходит путем восстановления контактирующей с ними воды. Это восстановление может обеспечиваться природным (ток воды), передвижением организмов в ее толще или за счет специальных дыхательных движений. С энергетической точки зрения пребывания в проточной воде - наиболее экономичное решение по восстановлению воды вокруг организма. Таким способом пользуются многие обитатели рек - реофильные черви, личинки поденок, веснянок, и другие организмы. Восстановление воды за счет поступательного движения самого организма осуществляется в ряду ракообразных, простейших, червей, личинок насекомых и многих других животных.
Смена воды у поглощающих кислород поверхностей за счет специальной работы организма встречается у представителей всех типов животных. Дыхательные движения, осуществляемые всем телом, свойственные многим червям, личинкам насекомых и некоторым рыбам, причем во многих случаях эти движения служат одновременно для фильтрации пищи. Например, многие полихет, живя в U-образных трубках, построенных в почве, волнообразными движениями прогоняют воду через свои норки, обеспечивает им дыхание и питание. Такое же двоякое значение имеет создание тока воды во многих личинок насекомых, погонофор, двустворчатых моллюсков, многих ракообразных и рыб. У ряда червей, личинок стрекоз и двукрылых со снижением содержания в воде кислорода темп дыхательных движений возрастает, что способствует поддержанию необходимого уровня газообмена. В тех случаях, когда вода в слое, где находится животное, крайне бедна кислородом и ее восстановление почти не улучшает условий дыхания, гидробионты приспосабливаются к забору воды из вышележащих, более аэрированных слоев. С этой целью двустворчатые моллюски извлекают вверх входные сифоны, личинки многих насекомых строят трубки, возвышающихся над грунтом.
Иногда восстановление воды достигается не созданием ее току, а движениями самого организма или его отдельных частей в толще воды. Так, например, колеблются, высунувшись из почвы задним концом тела много олигохет, в частности Tubifex tubifex, пиявок. Высунувшись вверх из норки и зацепившись за ее верхний край задним концом тела, качается рыбка трубочный угорь. Быстрые ритмичные движения жабрами делают многие рыбы и беспозвоночные. Чем хуже респираторные условия, тем оживленнее дыхательные движения.
Адаптация к использованию газообразного кислорода. Контакт дыхательных поверхностей с газообразным кислородом возможен в результате его захвата из атмосферы, с подледных скоплений, с воздухосодержащих тканей растений и пузырьков, образующихся в толще воды. Захват кислорода из атмосферы достигается либо путем периодического всплытия животных к поверхности воды, или выдвижением в воздух специальных дыхательных трубок. Всплытия для захвата кислорода встречается у некоторых насекомых и их личинок, у моллюсков, рыб и многих других животных. Всплывают к поверхности воды для дыхания легочные моллюски, земноводные и млекопитающие. Газообразным кислородом гидробионты могут дышать не только на поверхности воды, но и в ее толще. Так, паук Argyroneta aquatica строит под водой купол и заполняет его воздухом, принесенным с поверхности. Находясь в таком куполе, паук долгое время может не всплывать для дыхания. Многие насекомые, например жуки-плавунцы, в зимнее время, когда поверхность воды покрыта льдом, образуют на конце тела пузырек воздуха, который одевает дыхальца. Такой пузырек очень долго может обеспечивать дыхание животного, функционируя в качестве своеобразного «физического» жабры. По мере того как концентрация кислорода в пузырьке падает, газ начинает все сильнее диффундировать сюда из окружающей воды. Углекислота, наоборот, выводится из пузырька после того, как его концентрация начинает превышать ту, которая есть в воде.
Многие животные, находящиеся на дне, дышат атмосферным воздухом, не всплывая к поверхности и выставляя над водой дыхательную трубку. Иногда животные используют кислород, непосредственно выделяется растениями. Так, нимфы стрекоз могут заворачиваться в ковер из нитевидных водорослей и поглощать выделяемый ими кислород. Ряд животных использует для дыхания воздух, находящийся в тканях растений. Многие насекомые откладывают яйца в ткани растений, а воздух, содержащийся в них, обеспечивает дыхание зародышей. В некоторых случаях животные используют кислород, который выделяется симбиотическими водорослями. Таким образом, обеспечивают себе дыхание много простейших, кишечнополостных, червей, моллюсков.
Комбинирование водного и атмосферного дыхания. Способность к комбинированию воздушного и водного дыхания встречается у многих растений и животных, помогая им шире использовать те или другие респираторные ситуации. Комбинированный способ дыхания имеют, в частности, многие растения, листья которых плавают в воде. Последние, подобно листам наземных растений, несут устьица, но не на обеих сторонах, а только на верхней, причем их число очень велико. Например, в кувшинки Nymphaea alba оно достигает 400 на 1 мм, в рогоза - более 1300 на 1 мм. В наземных растений число устьиц меньше (в среднем 100-300 в 1 мм), так как площадь листовой поверхности, соприкасающейся с воздухом, больше и дыхания идет легче. Строение плавающих листьев такова, что предупреждает заливки устьиц водой во время волнений (соответствующая изогнутость листовой пластинки, восковой налет, что делает ее не смачиваемых).
Способность брать кислород из воды и с воздуха имеют много легочных моллюсков. Достаточно распространено комбинирование воздушного и водного дыхания у ракообразных. Личинки некоторых стрекоз при резком снижении концентрации кислорода в воде поднимаются к ее поверхности и забирают пузырек воздуха в заднюю кишку. Часто встречается комбинирование различных способов дыхания у рыб, например, в вьюнов, угрей и др.. В одних гидробионтов такая способность является адаптацией к существованию в среде, периодически или постоянно неблагоприятном в респираторном отношении. В других она обеспечивает возможность перехода из водной среды в воздушную для питания или расселения. Интересно отметить, что большинство гидробионтов, дышат растворенным и атмосферным кислородом, живет в тропической и субтропической областях, где высокая влажность воздуха облегчает животным выход на сушу.
Далеко не все гидробионты способны постоянно жить в бескислородной среде, то есть относятся к группе анаэробов (главным образом бактерии и простейшие). Подавляющее большинство обитателей водоемов нуждается
в кислороде, хотя некоторые из них, как было указано выше, могут иногда переносить его отсутствие и осуществлять аноксибиоз. Способность к нему в ряду гидробионтов-аэробов является адаптацией к перенесению неблагоприятных кислородных условий, периодически возникают в естественных местообитаниях.
В тех случаях, когда адаптации гидробионтов к существованию в условиях дефицита кислорода оказываются недостаточными, наступает гибель организмов. Если, вследствие резкого ухудшения кислородных условий в водоемах, приобретает массовый характер, то говорят о замор.
Способность гидробионтов выживать в воде с низкими концентрациями кислорода зависит от видовой принадлежности организмов, их состояния и условий внешней среды. Минимальная, или предельная концентрация кислорода, переносимая гидробионтами, как правило, ниже для организмов, обитающих в естественных местообитаниях в слабо аэрируемых воде. Поэтому пелагические формы обычно менее толерантны к низким концентрациям кислорода, чем бентосные, а среди последних жители ила выносливее форм, населяющих песок, глину или камни. По тем же причинам речные формы более требовательны к кислороду, чем озерные, так же как и холодноводные оксифильные обитатели быстротечных горных водоемов, нежели обитатели стоячих водоемов, прогреваемых сильнее. Величина предельной концентрации довольно сильно меняется с возрастом животных, обычно снижается у взрослых особей. Чувствительность к недостатку кислорода может обостряться на отдельных стадиях развития. Как правило, устойчивость к дефициту кислорода у животных разных систематических групп тем выше, чем они менее подвижны. Из внешних факторов на предельную концентрацию кислорода в наибольшей степени влияет температура. С ее повышением обмен организмов растет, их потребность в кислороде увеличивается и для ее удовлетворения нужны более благоприятные респираторные условия. Гидробионты имеют ряд биохимических адаптаций к обеспечению снабжения организма кислородом в условиях его дефицита. К ним относятся усиление вентиляции и кровообращения, повышение концентрации дыхательных пигментов, синтез их новых, более эффективных вариантов, изменение внутренней среды, что повышает способность пигментов связывать и освобождать кислород. Последняя адаптация (эффекты Бора и Рута) иногда проявляется в обратной форме - снижении чувствительности гемоглобина к повышению концентрации Н +, В некоторых активных рыб создаются очень высокие концентрации лактата, и рН может так падать, что затрудняет связывание кислорода гемоглобином в жабрах. У этих рыб найдены варианты гемоглобина, не чувствительны к Н +, т.е. не проявляют эффект Бора. Их обычно немного и они играют роль «аварийного резерва». При временной аноксии много гидробионтов могут осуществлять аэробный обмен за счет мобилизации запасов кислорода из каротиноидов, гем-содержащих пигментов и других депо. Обычно этих запасов хватает не более, чем на несколько десятков минут. Дальнейшее извлечения энергии может обеспечиваться во многих гидробионтов за счет анаэробного гликолиза и, вероятно, другими способами. Продолжительность нахождения и выживания гидробионтов при аноксии сильно варьирует, в зависимости от их видовой принадлежности, физиологического состояния и внешних условий. Формы, в которых лактат, образующийся при гликолизе, не накапливается, способны жить в отсутствие кислорода значительно дольше тех, которые накапливают молочную кислоту. Вторые, после перехода в аэробные условия, обнаруживают «екстрадихання» - повышение потребления кислорода, связано с окислением недоокисленных соединений, накопившиеся. В первых форм «екстрадихання» не наблюдается, «кислородной задолженности» не возникает и они при небольших энергетических затратах могут жить без кислорода в течение нескольких месяцев. Животные, накапливают при аноксибиози недоокисленные продукты, менее устойчивы к длительного отсутствия кислорода. Черепахи, погружаясь, могут жить за счет гликолиза несколько часов или дней и, всплывая, компенсируют «кислородную задолженность». Личинки хирономид хотя и накапливают лактат, но полностью теряют подвижность и находясь в состоянии пассивного анаэробиоза, выдерживают недели и месяцы аноксии. Устрицы и другие моллюски, замыкают раковины в отливной время, безболезненно выдерживают аноксию течение нескольких часов, накапливая продукты гликолиза. Способность к аноксибиозу наиболее характерна для представителей донной фауны, живущих в условиях периодического снижения концентрации кислорода до величин, близких к 0. При аноксибиозу они не питаются, теряют подвижность, перестают расти и развиваться. Концентрация кислорода, ниже которой гидробионты переходят от активного существования к пассивному, зависит от видовых особенностей, размера и других факторов. Как правило, формы, живущие в более аэрированная среде, впадают в аноксибиоз ранее, когда кислород присутствует еще в заметных количествах. Молодые организмы, обычно более требовательны к содержанию кислорода, исключаются из активной жизни ранее взрослых.
Заморы. В естественных водоемах нередко наблюдаются случаи массовой гибели гидробионтов от асфиксии. Они возникают не только вследствие дефицита кислорода, но и в результате накопления в воде значительных количеств углекислого газа, сероводорода и метана. Содержание этих газов обычно повышается параллельно снижению концентрации кислорода и потому особенно губителен для гидробионтов. При замора в первую очередь гибнут формы, менее устойчивы к недостатку кислорода, а затем и выносливые, до самых устойчивых, если катастрофическое ухудшение условий дыхания затягивается на долгий срок. В водоемах высоких широт заморы обычно возникают зимой, когда лед препятствует поступлению кислорода в воду из воздуха. Летние заморы обычно наблюдаются в стоячих водоемах, особенно во время массового появления водорослей. Днем результате фотосинтетической деятельности растений кислорода много, и ночью его концентрация резко снижается, и могут возникать заморные явления, сопровождающиеся гибелью животных.
Летние заморы имеют место не только в прудах и озерах, но даже в морях, например в Азовском и Балтийском. В Азовском море замор наблюдаются обычно с мая по август при тихой погоде, когда из-за отсутствия циркуляции воды содержание кислорода в толще, особенно у дна, падает до десятых долей миллиграмма в литре. Снижение концентрации кислорода у дна вызывается разложением водорослей, здесь отмирают. При замора в массе гибнет рыба и другие гидробионты, в частности моллюски. У берегов Перу раз в 11-12 лет происходит массовая гибель зоопланктона и рыб вследствие недостатка кислорода, когда сюда начинает подходить тепла экваториальная течение Эль-Ниньо.
Особенно остро протекают заморные явления зимой, когда они могут наблюдаться не только в стоячих водоемах, но даже и в реках. Например, грандиозные по своим масштабам ежегодные зимние заморы в г. Оби. Грунтовые воды, которыми она питается, содержат очень мало кислорода и много гуминовых веществ (сильно заболочена водосборная площадь). Когда после ледостава атмосферная аэрация воды практически прекращается, небольшие количества кислорода, находящегося в ней, быстро расходуются на окисление гуминовых кислот, и возникает замор. Обычно он начинается в конце декабря в верхнем течении реки и, распространяясь вниз со скоростью 30-40 км в сутки, достигает устья через 1,5-2 месяца. Прекращается замор в мае-июне, когда река начинает пополняться внешними водами. Содержание кислорода во время замора падает до 2-3% от нормального, и многие гидробионтов, особенно рыбы, погибают от удушья, хотя большая часть спасается в некоторых притоках Оби, где заморные явления не развиваются. Донные животные, более приспособлены к существованию в условиях дефицита кислорода, при заморе страдают меньше, чем пелагические.
В насыщенной кислородом воде содержание его не превышает 10 мл в 1 л, это в 21 раз ниже, чем в атмосфере. Поэтому условия дыхания гидробионтов значительно усложнены. Кислород поступает в воду в основном за счет фотосинтетической деятельности водорослей и диффузии из воздуха. Поэтому верхние слои водной толщи, как правило, богаче этим газом, чем нижние. С повышением температуры и солености воды концентрация в ней кислорода понижается. В слоях, сильно заселенных животными и бактериями, может создаваться резкий дефицит О2 из-за усиленного его потребления. Например, в Мировом океане богатые жизнью глубины от 50 до 1000 м характеризуются резким ухудшением аэрации – она в 7-10 раз ниже, чем в поверхностных водах, населенных фитопланктоном. Около дна водоемов условия могут быть близки к анаэробным.
Среди водных обитателей много видов, способных переносить широкие колебания содержания кислорода в воде, вплоть до почти полного его отсутствия (эвриоксибионты – «окси» – кислород, «бионт» – обитатель). К ним относятся, например, пресноводные олигохеты Tubifex tubifex, брюхоногие моллюски Viviparus viviparus. Среди рыб очень слабое насыщение воды кислородом могут выдерживать сазан, линь, караси. Рыба арапаима не имеет значительной конкуренции за жизненные ресурсы ввиду уникального механизма адаптации — арапайма может дышать воздухом из-за ткани, похожей на лёгочную ткань, которая выстилает глотку и плавательный пузырь.

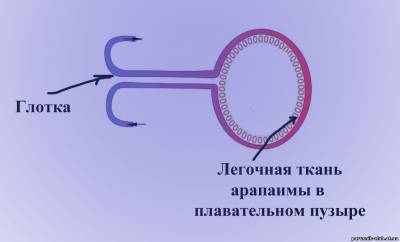
Рис. 1 Арапаима (слева) и ее дополнительный дыхательный аппарат в виде легочной ткани в плавательном пузыре - схема (справа).
Это приспособление развилось вследствие малого содержания кислорода в водах Амазонки. Таким образом, арапайма может переживать засуху, заглатывая воздух и зарываясь в грязь и песок болот.
У равноногих ракообразных свои уникальные способы адаптации различным, часто изменяющимся условиям кислородного режима. Особенно характерно строение брюшного отдела и его конечностей. Он всегда гораздо короче грудного, и количество составляющих его сегментов сокращено за счет того, что один, несколько или даже все сегменты срастаются с полукруглым или треугольным тельсоном, образуя крупный плеотельсон. Только у подотряда Anthuridea этого не происходит и последний брюшной сегмент и тельсон сохраняют самостоятельность. Конечности пяти передних брюшных сегментов состоят из короткого основания и двух широких листовидных, часто тонкостенных ветвей. Эти ветви накладываются друг на друга, как страницы книги, и направлены назад таким образом, что они находятся под плеотельсоном (рис. 2.). Основная функция передних брюшных ножек дыхательная. Они служат жабрами. Через их тонкие стенки растворенный в воде кислород проникает в проходящие в ножках кровеносные лакуны и окисляет кровь. В связи с этим, как уже указывалось, сердце у равноногих ракообразных в значительной степени смещено в брюшной отдел и расположено близко от жабр. Наружные ветви одной из пар брюшных ножек обычно сильно хитинизированы и служат крышкой, покрывающей остальные нижние жаберные листочки, которые, таким образом, оказываются сверху защищенными плеотельсоном, а снизу этой крышкой (рис. 2.). Такое строение дыхательного аппарата послужило, несомненно, предпосылкой, позволившей части равноногих перейти к сухопутному существованию. Их жабры оказались хорошо защищенными от высыхания и не потребовалось дополнительных усовершенствований для того, чтобы полностью их обезопасить в новых условиях существования. Большинство мокриц, несмотря на то что они живут на суше, дышат при помощи своих брюшных ножек, как настоящие водные животные: они используют кислород, растворенный в тонком слое покрывающей их жабры влаги. Следовательно, их жабры должны все время увлажняться. Это достигается у разных мокриц разными путями. У одних, как уже упоминалось, приток воды к жабрам обеспечивается благодаря своеобразной водопроводной системе, образованной скульптурой покровов. У других задние брюшные ножки — уроподы — устроены таким образом, что при соприкосновении с капелькой воды она, также под влиянием капиллярных сил, поступает в жабры по уроподам. Наконец, третьи орошают свои жабры жидкостью, выделяемой из анального отверстия и добываемой из поедаемой рачками сочной растительности. Однако есть мокрицы, способные при дыхании использовать не только растворенный в воде, но и атмосферный кислород. У таких мокриц в толще наружных ветвей передних брюшных ножек находится полость, соединяющаяся щелью с наружной средой, так что в эту полость свободно проникает воздух. От нее отходят ветвящиеся, заканчивающиеся слепо каналы, напоминающие трахеи насекомых и других наземных членистоногих и поэтому называемые псевдотрахеями (рис. 2.). Через них кислород воздуха проникает в кровь, циркулирующую в лакунах в толще наружных ветвей брюшных ножек, а кислород, растворенный в смачивающей жабры воде, используется внутренними ветвями.
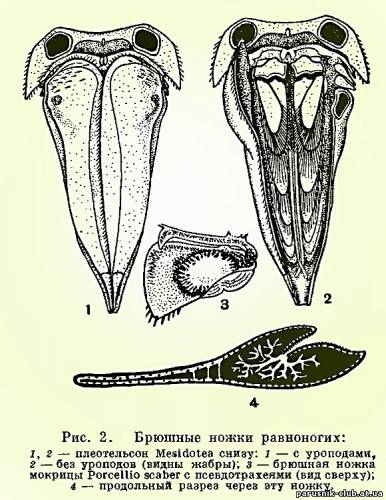
Рис. 2. Передние брюшные ножки ракообразных (мокриц).
Вместе с тем ряд видов стеноксибионтны – они могут существовать лишь при достаточно высоком насыщении воды кислородом (радужная форель, кумжа, гольян, ресничный червь Planaria alpina, личинки поденок, веснянок и др.). Многие виды способны при недостатке кислорода впадать в неактивное состояние – аноксибиоз – и таким образом переживать неблагоприятный период.
Дыхание гидробионтов осуществляется либо через поверхность тела, либо через специализированные органы – жабры, легкие, трахеи. При этом покровы могут служить дополнительным органом дыхания. Например, рыба вьюн через кожу потребляет в среднем до 63 % кислорода. Если через покровы тела происходит газообмен, то они очень тонки. Дыхание облегчается также увеличением поверхности. Это достигается в ходе эволюции видов образованием различных выростов, уплощением, удлинением, общим уменьшением размеров тела. Некоторые виды при недостатке кислорода активно изменяют величину дыхательной поверхности. Черви Tubifex tubifex сильно вытягивают тело в длину; гидры и актинии – щупальцы; иглокожие – амбулакральные ножки. Многие сидячие и малоподвижные животные обновляют вокруг себя воду, либо создавая ее направленный ток, либо колебательными движениями способствуя ее перемешиванию. Двустворчатым моллюскам для этой цели служат реснички, выстилающие стенки мантийной полости; ракообразным – работа брюшных или грудных ножек. Пиявки, личинки комаров-звонцов (мотыль), многие олигохеты колышут тело, высунувшись из грунта.
У некоторых видов встречается комбинирование водного и воздушного дыхания. Таковы двоякодышащие рыбы, сифонофоры дискофанты, многие легочные моллюски, ракообразные Gammarus lacustris и др. Вторичноводные животные сохраняют обычно атмосферный тип дыхания как более выгодный энергетически и нуждаются поэтому в контактах с воздушной средой, например ластоногие, китообразные, водяные жуки, личинки комаров и др.
Нехватка кислорода в воде приводит иногда к катастрофическим явлениям – заморам, сопровождающимся гибелью множества гидробионтов. Зимние заморы часто вызываются образованием на поверхности водоемов льда и прекращением контакта с воздухом; летние– повышением температуры воды и уменьшением вследствие этого растворимости кислорода.
Частая гибель рыб и многих беспозвоночных зимой характерна, например, для нижней части бассейна реки Оби, воды которой, стекающие из заболоченных пространств Западно-Сибирской низменности, крайне бедны растворенным кислородом. Иногда заморы возникают и в морях.
Кроме недостатка кислорода, заморы могут быть вызваны повышением концентрации в воде токсичных газов – метана, сероводорода, СО2 и др., образующихся в результате разложения органических материалов на дне водоемов.
Дыхание гидробионтов.
План.
1. Адаптация гидробионтов к газообмену.
2. Интенсивность газообмена.
3. Стойкость гидробионтов к дефициту кислорода.
4. Заморные явления.
Под интенсивностью дыхания понимается количество кислорода, потребляемого организмом в единицу времени на единицу массы. Количество потребленного кислорода оценивается в объемном или весовой единицах, вес выражается применительно ко всему веществу тела или его мягких частей (например, исключения веса раковины в моллюсков). Конечно, расчеты ведутся на сырую (живую) массу, реже на сухое вещество или его калорийный эквивалент. Отсутствие единой оценки количества вещества затрудняет анализ и сравнение имеющихся данных. Например, исключение из массы тяжелых скелетных образований при оценке дыхания моллюсков, усоногих раков и других завышает их обмен по сравнению с теми животными, у которых есть те же, но менее выраженные структуры, включают в общую массу (раковины рачков и панцири ракообразных, иглокожих и др..). Интенсивность дыхания организмов с различным содержанием воды сложно сравнивать. Поэтому необходимо проводить унификацию способов оценки массы организмов, количества вещества, на которую рассчитывается потребление кислорода. Большой интерес могут иметь данные об интенсивности дыхания, выраженные отношением энергетической стоимости кислорода, потребленного организмом, и энергоемкости последних. Такое отношение - хорошая характеристика метаболической активности живого вещества различных организмов и более точная мера интенсивности их газообмена.
Величина потребления кислорода характеризует уровень окислительных процессов, происходящих в организме, и, соответственно, его энергетические затраты в процессе жизнедеятельности. Сумма всех энергозатрат определяет величину обмена. Различают обмен основной, стандартный, активный и общий (средний). Под основным обменом, понимается уровень энергозатрат при полного покоя голодных организмов в условиях абиотической среды, близких к оптимальным. Стандартный обмен, близкий к основному, характеризует энергозатраты организмов с выключенной двигательной активностью в некоторых определенных условиях среды, в частности температурных. За величину активного обмена принимаются энергозатраты, связанные с обеспечением двигательной активности животных. Совокупность основного и активного обмена обозначают как общий или средний обмен. Потому что у беспозвоночных трудно не только вызвать состояние полного покоя, но и охарактеризовать это состояние, в опытах с гидробионтами обычно определяют стандартный или общий обмен. Животные содержатся в подопытных сосудах при отсутствии пищи тот или иной срок, после чего определяется суммарная величина потребляемого ими кислорода и вычисляется его среднее поглощения в единицу времени на единицу массы, т.е. средняя интенсивность дыхания.
Адаптация гидробионтов к газообмену.
Ускорение газообмена в гидробионтов, согласно законам диффузии может достигаться увеличением площади дыхательных поверхностей, истончением и разрыхлением покровов, через которые должны диффундировать газы
, созданием высокого градиента концентрации кислорода на внутренней и внешней сторонах покровов, в частности, путем приведения последних в контакт с аэрированная водой или газообразным кислородом.
Увеличение площади и газопроницаемости дыхательных поверхностей. Газообмен гидробионтов осуществляется либо через всю поверхность тела, или через его отдельные участки, превращены в специальные органы дыхания - жабры, трахеи, легкие и другие образования. Гидробионты, лишены специальных органов дыхания, обычно имеют тело с большой удельной поверхностью. Один из простейших способов ее увеличения заключается в уменьшении размера организмов. Небольшой размер характерен для простейших, коловраток, копепод, клещей и других организмов, не имеющих специальных органов дыхания. В мелких личинок комаров Chironоmus относительная величина жабр меньше, чем в крупных, удельная поверхность тела которых ниже, и на дыхательные органы ложится большая нагрузка. Икринки у рыб, живут и размножаются в озерах с пониженным содержанием кислорода, часто бывают мельче, чем у тех, которые живут и размножаются в озерах с более благоприятными условиями дыхания.
Увеличение поверхности тела часто достигается его уплощением, вытягиванием, образованием различных выростов, лопастей и т.п. С этим во многом связан, например, гидроморфоз растений, когда подводные листья расчленено значительно сильнее надводного, что находится в более благоприятных для дыхания условиях. Степень развития дыхательных поверхностей часто обнаруживает тесную зависимость от респираторных условий. У крабов, часто находятся на воздухе, где кислорода больше, наблюдается редукция числа и величины жабр. Иногда животные сами активно увеличивают дыхательную поверхность за счет изменения формы тела. Например, олигохеты в неблагоприятных условиях дыхания, сильно вытягиваясь в длину, утончаются, благодаря чему поверхность их тела увеличивается. Гидры и актинии при недостатке кислорода сильно вытягивают свое тело и щупальца, иглокожие - амбулакральные ножки.
Скорость диффузии газов определяется не только величиной дыхательной поверхности, но и толщиной покровов, через которые происходит газообмен. В силу этого истончение покровов дыхательных поверхностей представляет собой одну из основных адаптации к газообмена. Очень тонкие покровы на жабрах, в легких и других специальных образованиях, которые функционируют как органы дыхания. Если специальных органов дыхания нет, то утончаются покровы всего тела. Некоторые гидробионты способны увеличивать проницаемость наружных покровов, когда возникает необходимость усиления газообмена. Так, в соответствии с потребностями газообмена, изменяется степень проницаемости оболочек в икринок рыб за счет выделения гиалуронидазы.
Адаптация к использованию растворенного кислорода. Контакт дыхательных поверхностей с хорошо аэрированных водой, прежде всего, достигается в гидробионтов выбором местонахождения, где вода имеет достаточно высокое содержание кислорода, а также ее периодическим обновлением вблизи своего тела. В случае резкого ухудшения респираторного среды во организмов перемещаются даже в не свойственные им биотопы. Например, сувойка Vorticella nebulifera при дефиците кислорода образует заднее кольцо ресничек, отделяется от стебелька и ведет планктонный образ жизни до тех пор, пока респираторные условия у дна не изменятся на лучшие. Черви Nereis, личинки комара Chironomus и ряд других животных выползают на поверхность почвы, когда респираторные условия в его толще резко ухудшаются. Выбор мест с более благоприятными условиями дыхания наблюдается и в пелагических животных, когда они оставляют участки водоема с недостаточной для них концентрацией кислорода. Особенно ярко это явление наблюдается при замора, когда рыбы, клопы, жуки и другие животные нередко перемещаются на значительные расстояния в поисках достаточно аэрируемой воды.
Универсальная адаптация гидробионтов к аэрированию дыхательных поверхностей происходит путем восстановления контактирующей с ними воды. Это восстановление может обеспечиваться природным (ток воды), передвижением организмов в ее толще или за счет специальных дыхательных движений. С энергетической точки зрения пребывания в проточной воде - наиболее экономичное решение по восстановлению воды вокруг организма. Таким способом пользуются многие обитатели рек - реофильные черви, личинки поденок, веснянок, и другие организмы. Восстановление воды за счет поступательного движения самого организма осуществляется в ряду ракообразных, простейших, червей, личинок насекомых и многих других животных.
Смена воды у поглощающих кислород поверхностей за счет специальной работы организма встречается у представителей всех типов животных. Дыхательные движения, осуществляемые всем телом, свойственные многим червям, личинкам насекомых и некоторым рыбам, причем во многих случаях эти движения служат одновременно для фильтрации пищи. Например, многие полихет, живя в U-образных трубках, построенных в почве, волнообразными движениями прогоняют воду через свои норки, обеспечивает им дыхание и питание. Такое же двоякое значение имеет создание тока воды во многих личинок насекомых, погонофор, двустворчатых моллюсков, многих ракообразных и рыб. У ряда червей, личинок стрекоз и двукрылых со снижением содержания в воде кислорода темп дыхательных движений возрастает, что способствует поддержанию необходимого уровня газообмена. В тех случаях, когда вода в слое, где находится животное, крайне бедна кислородом и ее восстановление почти не улучшает условий дыхания, гидробионты приспосабливаются к забору воды из вышележащих, более аэрированных слоев. С этой целью двустворчатые моллюски извлекают вверх входные сифоны, личинки многих насекомых строят трубки, возвышающихся над грунтом.
Иногда восстановление воды достигается не созданием ее току, а движениями самого организма или его отдельных частей в толще воды. Так, например, колеблются, высунувшись из почвы задним концом тела много олигохет, в частности Tubifex tubifex, пиявок. Высунувшись вверх из норки и зацепившись за ее верхний край задним концом тела, качается рыбка трубочный угорь. Быстрые ритмичные движения жабрами делают многие рыбы и беспозвоночные. Чем хуже респираторные условия, тем оживленнее дыхательные движения.
Адаптация к использованию газообразного кислорода. Контакт дыхательных поверхностей с газообразным кислородом возможен в результате его захвата из атмосферы, с подледных скоплений, с воздухосодержащих тканей растений и пузырьков, образующихся в толще воды. Захват кислорода из атмосферы достигается либо путем периодического всплытия животных к поверхности воды, или выдвижением в воздух специальных дыхательных трубок. Всплытия для захвата кислорода встречается у некоторых насекомых и их личинок, у моллюсков, рыб и многих других животных. Всплывают к поверхности воды для дыхания легочные моллюски, земноводные и млекопитающие. Газообразным кислородом гидробионты могут дышать не только на поверхности воды, но и в ее толще. Так, паук Argyroneta aquatica строит под водой купол и заполняет его воздухом, принесенным с поверхности. Находясь в таком куполе, паук долгое время может не всплывать для дыхания. Многие насекомые, например жуки-плавунцы, в зимнее время, когда поверхность воды покрыта льдом, образуют на конце тела пузырек воздуха, который одевает дыхальца. Такой пузырек очень долго может обеспечивать дыхание животного, функционируя в качестве своеобразного «физического» жабры. По мере того как концентрация кислорода в пузырьке падает, газ начинает все сильнее диффундировать сюда из окружающей воды. Углекислота, наоборот, выводится из пузырька после того, как его концентрация начинает превышать ту, которая есть в воде.
Многие животные, находящиеся на дне, дышат атмосферным воздухом, не всплывая к поверхности и выставляя над водой дыхательную трубку. Иногда животные используют кислород, непосредственно выделяется растениями. Так, нимфы стрекоз могут заворачиваться в ковер из нитевидных водорослей и поглощать выделяемый ими кислород. Ряд животных использует для дыхания воздух, находящийся в тканях растений. Многие насекомые откладывают яйца в ткани растений, а воздух, содержащийся в них, обеспечивает дыхание зародышей. В некоторых случаях животные используют кислород, который выделяется симбиотическими водорослями. Таким образом, обеспечивают себе дыхание много простейших, кишечнополостных, червей, моллюсков.
Комбинирование водного и атмосферного дыхания. Способность к комбинированию воздушного и водного дыхания встречается у многих растений и животных, помогая им шире использовать те или другие респираторные ситуации. Комбинированный способ дыхания имеют, в частности, многие растения, листья которых плавают в воде. Последние, подобно листам наземных растений, несут устьица, но не на обеих сторонах, а только на верхней, причем их число очень велико. Например, в кувшинки Nymphaea alba оно достигает 400 на 1 мм, в рогоза - более 1300 на 1 мм. В наземных растений число устьиц меньше (в среднем 100-300 в 1 мм), так как площадь листовой поверхности, соприкасающейся с воздухом, больше и дыхания идет легче. Строение плавающих листьев такова, что предупреждает заливки устьиц водой во время волнений (соответствующая изогнутость листовой пластинки, восковой налет, что делает ее не смачиваемых).
Способность брать кислород из воды и с воздуха имеют много легочных моллюсков. Достаточно распространено комбинирование воздушного и водного дыхания у ракообразных. Личинки некоторых стрекоз при резком снижении концентрации кислорода в воде поднимаются к ее поверхности и забирают пузырек воздуха в заднюю кишку. Часто встречается комбинирование различных способов дыхания у рыб, например, в вьюнов, угрей и др.. В одних гидробионтов такая способность является адаптацией к существованию в среде, периодически или постоянно неблагоприятном в респираторном отношении. В других она обеспечивает возможность перехода из водной среды в воздушную для питания или расселения. Интересно отметить, что большинство гидробионтов, дышат растворенным и атмосферным кислородом, живет в тропической и субтропической областях, где высокая влажность воздуха облегчает животным выход на сушу.
Далеко не все гидробионты способны постоянно жить в бескислородной среде, то есть относятся к группе анаэробов (главным образом бактерии и простейшие). Подавляющее большинство обитателей водоемов нуждается
в кислороде, хотя некоторые из них, как было указано выше, могут иногда переносить его отсутствие и осуществлять аноксибиоз. Способность к нему в ряду гидробионтов-аэробов является адаптацией к перенесению неблагоприятных кислородных условий, периодически возникают в естественных местообитаниях.
В тех случаях, когда адаптации гидробионтов к существованию в условиях дефицита кислорода оказываются недостаточными, наступает гибель организмов. Если, вследствие резкого ухудшения кислородных условий в водоемах, приобретает массовый характер, то говорят о замор.
Способность гидробионтов выживать в воде с низкими концентрациями кислорода зависит от видовой принадлежности организмов, их состояния и условий внешней среды. Минимальная, или предельная концентрация кислорода, переносимая гидробионтами, как правило, ниже для организмов, обитающих в естественных местообитаниях в слабо аэрируемых воде. Поэтому пелагические формы обычно менее толерантны к низким концентрациям кислорода, чем бентосные, а среди последних жители ила выносливее форм, населяющих песок, глину или камни. По тем же причинам речные формы более требовательны к кислороду, чем озерные, так же как и холодноводные оксифильные обитатели быстротечных горных водоемов, нежели обитатели стоячих водоемов, прогреваемых сильнее. Величина предельной концентрации довольно сильно меняется с возрастом животных, обычно снижается у взрослых особей. Чувствительность к недостатку кислорода может обостряться на отдельных стадиях развития. Как правило, устойчивость к дефициту кислорода у животных разных систематических групп тем выше, чем они менее подвижны. Из внешних факторов на предельную концентрацию кислорода в наибольшей степени влияет температура. С ее повышением обмен организмов растет, их потребность в кислороде увеличивается и для ее удовлетворения нужны более благоприятные респираторные условия. Гидробионты имеют ряд биохимических адаптаций к обеспечению снабжения организма кислородом в условиях его дефицита. К ним относятся усиление вентиляции и кровообращения, повышение концентрации дыхательных пигментов, синтез их новых, более эффективных вариантов, изменение внутренней среды, что повышает способность пигментов связывать и освобождать кислород. Последняя адаптация (эффекты Бора и Рута) иногда проявляется в обратной форме - снижении чувствительности гемоглобина к повышению концентрации Н +, В некоторых активных рыб создаются очень высокие концентрации лактата, и рН может так падать, что затрудняет связывание кислорода гемоглобином в жабрах. У этих рыб найдены варианты гемоглобина, не чувствительны к Н +, т.е. не проявляют эффект Бора. Их обычно немного и они играют роль «аварийного резерва». При временной аноксии много гидробионтов могут осуществлять аэробный обмен за счет мобилизации запасов кислорода из каротиноидов, гем-содержащих пигментов и других депо. Обычно этих запасов хватает не более, чем на несколько десятков минут. Дальнейшее извлечения энергии может обеспечиваться во многих гидробионтов за счет анаэробного гликолиза и, вероятно, другими способами. Продолжительность нахождения и выживания гидробионтов при аноксии сильно варьирует, в зависимости от их видовой принадлежности, физиологического состояния и внешних условий. Формы, в которых лактат, образующийся при гликолизе, не накапливается, способны жить в отсутствие кислорода значительно дольше тех, которые накапливают молочную кислоту. Вторые, после перехода в аэробные условия, обнаруживают «екстрадихання» - повышение потребления кислорода, связано с окислением недоокисленных соединений, накопившиеся. В первых форм «екстрадихання» не наблюдается, «кислородной задолженности» не возникает и они при небольших энергетических затратах могут жить без кислорода в течение нескольких месяцев. Животные, накапливают при аноксибиози недоокисленные продукты, менее устойчивы к длительного отсутствия кислорода. Черепахи, погружаясь, могут жить за счет гликолиза несколько часов или дней и, всплывая, компенсируют «кислородную задолженность». Личинки хирономид хотя и накапливают лактат, но полностью теряют подвижность и находясь в состоянии пассивного анаэробиоза, выдерживают недели и месяцы аноксии. Устрицы и другие моллюски, замыкают раковины в отливной время, безболезненно выдерживают аноксию течение нескольких часов, накапливая продукты гликолиза. Способность к аноксибиозу наиболее характерна для представителей донной фауны, живущих в условиях периодического снижения концентрации кислорода до величин, близких к 0. При аноксибиозу они не питаются, теряют подвижность, перестают расти и развиваться. Концентрация кислорода, ниже которой гидробионты переходят от активного существования к пассивному, зависит от видовых особенностей, размера и других факторов. Как правило, формы, живущие в более аэрированная среде, впадают в аноксибиоз ранее, когда кислород присутствует еще в заметных количествах. Молодые организмы, обычно более требовательны к содержанию кислорода, исключаются из активной жизни ранее взрослых.
Заморы. В естественных водоемах нередко наблюдаются случаи массовой гибели гидробионтов от асфиксии. Они возникают не только вследствие дефицита кислорода, но и в результате накопления в воде значительных количеств углекислого газа, сероводорода и метана. Содержание этих газов обычно повышается параллельно снижению концентрации кислорода и потому особенно губителен для гидробионтов. При замора в первую очередь гибнут формы, менее устойчивы к недостатку кислорода, а затем и выносливые, до самых устойчивых, если катастрофическое ухудшение условий дыхания затягивается на долгий срок. В водоемах высоких широт заморы обычно возникают зимой, когда лед препятствует поступлению кислорода в воду из воздуха. Летние заморы обычно наблюдаются в стоячих водоемах, особенно во время массового появления водорослей. Днем результате фотосинтетической деятельности растений кислорода много, и ночью его концентрация резко снижается, и могут возникать заморные явления, сопровождающиеся гибелью животных.
Летние заморы имеют место не только в прудах и озерах, но даже в морях, например в Азовском и Балтийском. В Азовском море замор наблюдаются обычно с мая по август при тихой погоде, когда из-за отсутствия циркуляции воды содержание кислорода в толще, особенно у дна, падает до десятых долей миллиграмма в литре. Снижение концентрации кислорода у дна вызывается разложением водорослей, здесь отмирают. При замора в массе гибнет рыба и другие гидробионты, в частности моллюски. У берегов Перу раз в 11-12 лет происходит массовая гибель зоопланктона и рыб вследствие недостатка кислорода, когда сюда начинает подходить тепла экваториальная течение Эль-Ниньо.
Особенно остро протекают заморные явления зимой, когда они могут наблюдаться не только в стоячих водоемах, но даже и в реках. Например, грандиозные по своим масштабам ежегодные зимние заморы в г. Оби. Грунтовые воды, которыми она питается, содержат очень мало кислорода и много гуминовых веществ (сильно заболочена водосборная площадь). Когда после ледостава атмосферная аэрация воды практически прекращается, небольшие количества кислорода, находящегося в ней, быстро расходуются на окисление гуминовых кислот, и возникает замор. Обычно он начинается в конце декабря в верхнем течении реки и, распространяясь вниз со скоростью 30-40 км в сутки, достигает устья через 1,5-2 месяца. Прекращается замор в мае-июне, когда река начинает пополняться внешними водами. Содержание кислорода во время замора падает до 2-3% от нормального, и многие гидробионтов, особенно рыбы, погибают от удушья, хотя большая часть спасается в некоторых притоках Оби, где заморные явления не развиваются. Донные животные, более приспособлены к существованию в условиях дефицита кислорода, при заморе страдают меньше, чем пелагические.
В насыщенной кислородом воде содержание его не превышает 10 мл в 1 л, это в 21 раз ниже, чем в атмосфере. Поэтому условия дыхания гидробионтов значительно усложнены. Кислород поступает в воду в основном за счет фотосинтетической деятельности водорослей и диффузии из воздуха. Поэтому верхние слои водной толщи, как правило, богаче этим газом, чем нижние. С повышением температуры и солености воды концентрация в ней кислорода понижается. В слоях, сильно заселенных животными и бактериями, может создаваться резкий дефицит О2 из-за усиленного его потребления. Например, в Мировом океане богатые жизнью глубины от 50 до 1000 м характеризуются резким ухудшением аэрации – она в 7-10 раз ниже, чем в поверхностных водах, населенных фитопланктоном. Около дна водоемов условия могут быть близки к анаэробным.
Среди водных обитателей много видов, способных переносить широкие колебания содержания кислорода в воде, вплоть до почти полного его отсутствия (эвриоксибионты – «окси» – кислород, «бионт» – обитатель). К ним относятся, например, пресноводные олигохеты Tubifex tubifex, брюхоногие моллюски Viviparus viviparus. Среди рыб очень слабое насыщение воды кислородом могут выдерживать сазан, линь, караси. Рыба арапаима не имеет значительной конкуренции за жизненные ресурсы ввиду уникального механизма адаптации — арапайма может дышать воздухом из-за ткани, похожей на лёгочную ткань, которая выстилает глотку и плавательный пузырь.


Это приспособление развилось вследствие малого содержания кислорода в водах Амазонки. Таким образом, арапайма может переживать засуху, заглатывая воздух и зарываясь в грязь и песок болот.
У равноногих ракообразных свои уникальные способы адаптации различным, часто изменяющимся условиям кислородного режима. Особенно характерно строение брюшного отдела и его конечностей. Он всегда гораздо короче грудного, и количество составляющих его сегментов сокращено за счет того, что один, несколько или даже все сегменты срастаются с полукруглым или треугольным тельсоном, образуя крупный плеотельсон. Только у подотряда Anthuridea этого не происходит и последний брюшной сегмент и тельсон сохраняют самостоятельность. Конечности пяти передних брюшных сегментов состоят из короткого основания и двух широких листовидных, часто тонкостенных ветвей. Эти ветви накладываются друг на друга, как страницы книги, и направлены назад таким образом, что они находятся под плеотельсоном (рис. 2.). Основная функция передних брюшных ножек дыхательная. Они служат жабрами. Через их тонкие стенки растворенный в воде кислород проникает в проходящие в ножках кровеносные лакуны и окисляет кровь. В связи с этим, как уже указывалось, сердце у равноногих ракообразных в значительной степени смещено в брюшной отдел и расположено близко от жабр. Наружные ветви одной из пар брюшных ножек обычно сильно хитинизированы и служат крышкой, покрывающей остальные нижние жаберные листочки, которые, таким образом, оказываются сверху защищенными плеотельсоном, а снизу этой крышкой (рис. 2.). Такое строение дыхательного аппарата послужило, несомненно, предпосылкой, позволившей части равноногих перейти к сухопутному существованию. Их жабры оказались хорошо защищенными от высыхания и не потребовалось дополнительных усовершенствований для того, чтобы полностью их обезопасить в новых условиях существования. Большинство мокриц, несмотря на то что они живут на суше, дышат при помощи своих брюшных ножек, как настоящие водные животные: они используют кислород, растворенный в тонком слое покрывающей их жабры влаги. Следовательно, их жабры должны все время увлажняться. Это достигается у разных мокриц разными путями. У одних, как уже упоминалось, приток воды к жабрам обеспечивается благодаря своеобразной водопроводной системе, образованной скульптурой покровов. У других задние брюшные ножки — уроподы — устроены таким образом, что при соприкосновении с капелькой воды она, также под влиянием капиллярных сил, поступает в жабры по уроподам. Наконец, третьи орошают свои жабры жидкостью, выделяемой из анального отверстия и добываемой из поедаемой рачками сочной растительности. Однако есть мокрицы, способные при дыхании использовать не только растворенный в воде, но и атмосферный кислород. У таких мокриц в толще наружных ветвей передних брюшных ножек находится полость, соединяющаяся щелью с наружной средой, так что в эту полость свободно проникает воздух. От нее отходят ветвящиеся, заканчивающиеся слепо каналы, напоминающие трахеи насекомых и других наземных членистоногих и поэтому называемые псевдотрахеями (рис. 2.). Через них кислород воздуха проникает в кровь, циркулирующую в лакунах в толще наружных ветвей брюшных ножек, а кислород, растворенный в смачивающей жабры воде, используется внутренними ветвями.
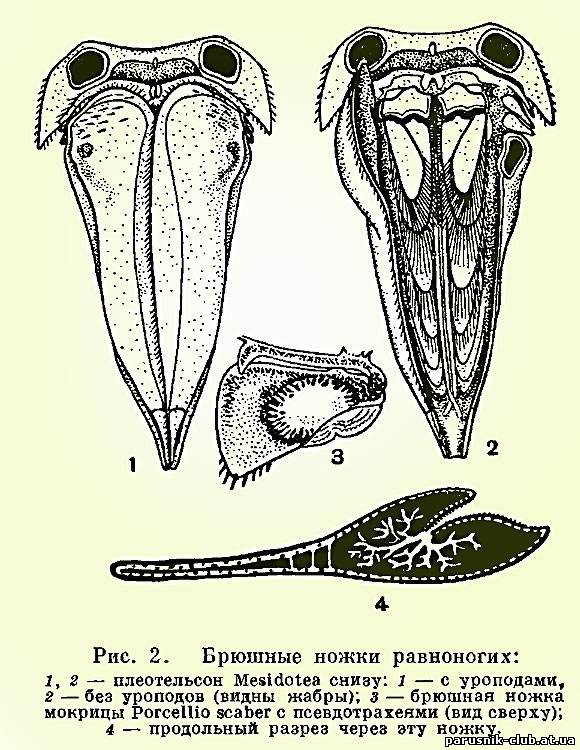
Вместе с тем ряд видов стеноксибионтны – они могут существовать лишь при достаточно высоком насыщении воды кислородом (радужная форель, кумжа, гольян, ресничный червь Planaria alpina, личинки поденок, веснянок и др.). Многие виды способны при недостатке кислорода впадать в неактивное состояние – аноксибиоз – и таким образом переживать неблагоприятный период.
Дыхание гидробионтов осуществляется либо через поверхность тела, либо через специализированные органы – жабры, легкие, трахеи. При этом покровы могут служить дополнительным органом дыхания. Например, рыба вьюн через кожу потребляет в среднем до 63 % кислорода. Если через покровы тела происходит газообмен, то они очень тонки. Дыхание облегчается также увеличением поверхности. Это достигается в ходе эволюции видов образованием различных выростов, уплощением, удлинением, общим уменьшением размеров тела. Некоторые виды при недостатке кислорода активно изменяют величину дыхательной поверхности. Черви Tubifex tubifex сильно вытягивают тело в длину; гидры и актинии – щупальцы; иглокожие – амбулакральные ножки. Многие сидячие и малоподвижные животные обновляют вокруг себя воду, либо создавая ее направленный ток, либо колебательными движениями способствуя ее перемешиванию. Двустворчатым моллюскам для этой цели служат реснички, выстилающие стенки мантийной полости; ракообразным – работа брюшных или грудных ножек. Пиявки, личинки комаров-звонцов (мотыль), многие олигохеты колышут тело, высунувшись из грунта.
У некоторых видов встречается комбинирование водного и воздушного дыхания. Таковы двоякодышащие рыбы, сифонофоры дискофанты, многие легочные моллюски, ракообразные Gammarus lacustris и др. Вторичноводные животные сохраняют обычно атмосферный тип дыхания как более выгодный энергетически и нуждаются поэтому в контактах с воздушной средой, например ластоногие, китообразные, водяные жуки, личинки комаров и др.
Нехватка кислорода в воде приводит иногда к катастрофическим явлениям – заморам, сопровождающимся гибелью множества гидробионтов. Зимние заморы часто вызываются образованием на поверхности водоемов льда и прекращением контакта с воздухом; летние– повышением температуры воды и уменьшением вследствие этого растворимости кислорода.
Частая гибель рыб и многих беспозвоночных зимой характерна, например, для нижней части бассейна реки Оби, воды которой, стекающие из заболоченных пространств Западно-Сибирской низменности, крайне бедны растворенным кислородом. Иногда заморы возникают и в морях.
Кроме недостатка кислорода, заморы могут быть вызваны повышением концентрации в воде токсичных газов – метана, сероводорода, СО2 и др., образующихся в результате разложения органических материалов на дне водоемов.






